Autor: ventero
viernes, 01 de junio de 2007
Sección: Artículos generales
Información publicada por: ventero
Mostrado 59.409 veces.
Los ancestros del hombre comenzaron a caminar primero en los árboles
Los primates que antecedieron al hombre moderno comenzaron a caminar en los árboles antes de hacerlo en suelo firme, según afirma un estudio de paleontólogos ingleses divulgado este jueves por la revista 'Science'.
De "El Mundo", Madrid, y EFE, Washington.-
Los primates que antecedieron al hombre moderno comenzaron a caminar en los árboles antes de hacerlo en suelo firme, según afirma un estudio de paleontólogos ingleses divulgado el 31 de mayo de 2007 por la revista 'Science'.

Las actuales teorías sobre la evolución de los homínidos señalaban que el bipedalismo, la característica que diferencia al hombre de sus parientes primates, comenzó cuando éstos descendieron de los árboles, no antes. Y una de las explicaciones más socorridas ha sido que los ancestros de chimpancés, gorilas y, por último, del hombre descendieron de los árboles para desplazarse en cuatro patas. Con el tiempo, comenzaron a caminar apoyándose en los nudillos y después únicamente sobre sus extremidades inferiores, como los seres humanos.
Sin embargo, según los paleontólogos de la Universidad de Liverpool, ese razonamiento comenzó a complicarse cuando estudios recientes hechos a partir de fósiles mostraron que algunos homínidos, incluido el de Lucy (australopithecus afarensis), vivieron en bosques.

Al mismo tiempo, otras formas más primitivas como la del Hombre del Milenio (Orrorin) parece que vivieron en la copa de los árboles y se desplazaban sobre sus extremidades inferiores. "Los resultados de nuestro estudio han confundido el panorama", señaló Robin Crompton, uno de los autores de la investigación. "Si estamos en lo cierto, esto significa que no se puede confiar en que el bipedalismo nos diga si estamos ante el ancestro de un ser humano o el de otro primate", indicó. Según el científico, "es cada vez más difícil decir qué es humano y qué es simio y nuestro trabajo lo confirma".
Crompton, Susannah Thorpe y Roger Holder, de la Universidad de Birmingham, llegaron a la conclusión sobre el bipedalismo arbóreo de los primates al observar orangutanes en Sumatra (Indonesia). Según señalaron, los orangutanes pasan la mayor parte de su vida en los árboles, lo que los convierte en modelos útiles para establecer cómo se desplazaron nuestros ancestros hace millones de años.
Debido a que esos ancestros comían frutas, habrían tenido que apoyarse en las ramas débiles en la periferia de la copa de los árboles. El hecho de caminar sobre sus extremidades inferiores y utilizar sus brazos para mantener el equilibrio les habría ayudado a desplazarse, según el estudio. "Nuestros resultados sugieren que el bipedalismo se usó para recorrer las ramas más débiles, donde están las mejores frutas, y también para pasar de un árbol a otro", según Thorpe.
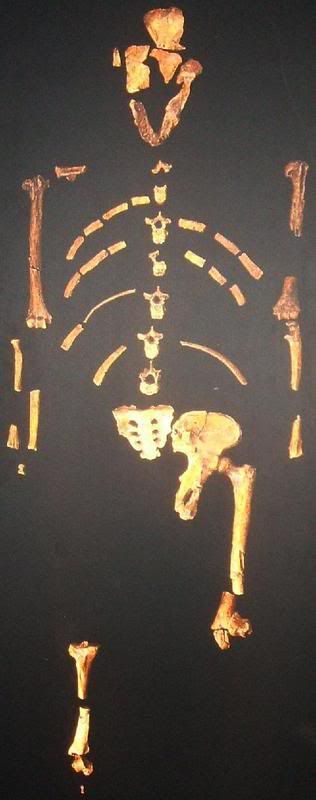
Los científicos creen que al bipedalismo contribuyó un cambio climático registrado entre 5 y 24 millones de años atrás, cuando disminuyó la frondosidad de los árboles. En un ambiente más árido, comenzaron a encontrar dificultades para pasar de un árbol a otro debido a la disminución de la espesura. Los ancestros humanos respondieron a este problema abandonando las copas de los árboles para llegar al suelo de las selvas, donde mantuvieron su bipedalismo y comenzaron a alimentarse en árboles más pequeños. Por su parte, los ancestros de chimpancés y gorilas se especializaron más en el ascenso vertical de los árboles y desarrollaron su forma de caminar sobre los nudillos.
"Nuestra conclusión es que el bipedalismo arbóreo tuvo grandes beneficios adaptativos. Por ello no necesitamos explicar cómo nuestros ancestros pasaron de ser cuadrúpedos a bípedos", dijo Thorpe.
-
No hay imágenes relacionadas.
Comentarios cerrados por el administrador Silberius
Motivo: Degradación en la calidad de los comentarios
Comentarios
 Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Tu único clic no la borarrá, pero contribuirá a que la sabiduría del grupo pueda funcionar correctamente.
Comentarios cerrados por el administrador Silberius
Motivo: Degradación en la calidad de los comentarios
Brujo, no se preocupe usted que ya andaba yo vacunado sobre este virus!
ya me inmunice con un tal Kronos....!
Saludos
Desde luego, es flipante la gente con la que se puede topar uno en un foro. Encomiables los esfuerzos de Etpoursimouve y el Brujo Redivivo por explicar conceptos básicos de la ciencia a unos fanáticos religiosos. Sin embargo, a tenor del talante que han demostrado, me temo que sea inútil.
Me parece que el filósofo Michael Ruse achaca el auge del posmodernismo, entre otras cosas, al clima de desconfianza hacia la ciencia que crearon los acontecimientos de la Segunda Guerra Mundial y la subsiguiente guerra fría. Aparte, como alguien ha apuntado más arriba, se trata de un problema de capacidad y esfuerzo intelectual. Es mucho más fácil entender lo que dice el Corán que una teoría científica medianamente compleja.
Lo que me fastidia es que generaciones de científicos dediquen su vida a trabajar dura y honradamente para intentar esclarecer hechos importantes de la realidad, y que haya gente que piense que puede desacreditarlos tranquilamente a base de citas de un libro de fantasía. Eso, cuando no los queman en una hoguera.
Respecto a la pregunta de Apomios sobre si sé lo que es el gen de la herencia, sólo le contestaré que soy biólogo.
En cuanto al debate en sí del bipedismo, entiendo que, según nos cuenta Etpoursimouve, la hipótesis es que los primeros homínidos vivían en ambientes boscosos. Eso es distinto, muy distinto diría yo, a que fueran arborícolas. Simplemente, no me puedo imaginar que un animal de cierto tamaño pudiera moverse andando por los árboles con un mínimo de soltura. Y desde luego, no mejor que un cuadrúpedo, con lo que la marcha erguida no tendría ninguna ventaja adaptativa, sino más bien al contrario.
Puedo concebir que el bipedismo evolucionara en las selvas, pero no que lo hiciera en los árboles.
El cambio de la marcha cuadrúpeda a un bipedismo eficaz implica, entre otras cosas:
-El desplazamiento del foramen magnum de la parte trasera del cráneo a la base del mismo.
-El desplazamiento de las ramas ilíacas de la pelvis hacia atrás, con aumento de anchura para permitir la inserción de los glúteos, músculos importantes para el equilibrio.
-Pérdida de curvatura del fémur, que se hace más rectilíneo.
-Cambio en la morfología del pie, aparición del arco plantar y desaparición del pulgar oponible.
Son muchos e importantes cambios. Ninguno de ellos es, en principio, útil en una vida arborícola, por lo que estarían seleccionados negativamente. Lo más lógico es que evolucionaran en un animal que viviera ya sobre el suelo, aunque fuera en una selva.
Un saludo.
Tranquilo, Brigantinus, que las personas medianamente inteligentes ya saben que el El Brujo y yo no somos la misma persona, aunque te agradezco la aclaración, por si algún ingenuo hiciera caso de determinadas tontadas. Lo único es que, últimamente, se nos ve juntos en el, no negaré que divertido a ratos, empeño de dejar con el culo al aire (cosa fácil, por cierto) a algunos creacionistas, fanáticos religiosos, inventores de historias, abrazapiedras varios y, en fin, por no extenderme, mentirosos en general.
Lo que es encomiable de verdad es ver los ímprobos esfuerzos de Ramonmo, Servan y etpoursimouve, entre otros, por poner las cosas en su sitio. No desesperen que, aunque a alguien como Apomios no le van a convencer con argumentos, muchos estamos aprendiendo bastante sobre este tema. Aunque sólo sea por seo, creo que merecen la pena sus esfuerzos.
Vaya Brujo, parece que las huestes creacionistas se ponen nerviosos. Eso sí, vaya aliados solventes se han buscado.
Saludos
Ramonmo, solo decirte que los Orangutanes pueden pesar hasta 80 Kg, y sin embargo siguen siendo arborícolas. Pero yo no intento colar la idea de que todos los rasgos que se identifican con el bipedismo actual, asi como lo que muy acertadamente has descrito sobre los cambios anatómicos, fueran desarrollados encima de un árbol. Eso tengo claro que se desarrollo debajo y no encima. Solo apunto esa posibilidad y la duda de lo que la cuestión es, y lo qué subyace en el desarrollo de la marcha erguida en espacios cerrados? en cualquier caso no podemos vaticinar nada, así que serán los que están investigando los que nos sacaran de la eterna cuestión en torno al bipedismo. Lo bueno es que al final todos nos replanteamos aspectos que se creian dogmas,
y como la cosa va de citas ( al menos por mi parte), citare una mas afortunada de un profesor de Invertebrados de 4°curso: "nada es tajante en Biología, ya que es la ciencia de las excepciones"
Y esto si que nos deja en un punto muerto! esperando la labor de los investigadores....
Un saludo fraternal Ramonmo, y su entera disposición
Apomios05 de jun. 2007Seras biologo de alguna religion radical, pq defender teorias como un dogma de fe, es eso mismo.
EL IMPACTO DE LAMARCK
Ahora bien, ¿cómo pueden ocurrir estas “variaciones favorables”? Darwin trató de responder a esta cuestión partiendo del punto de vista que el primitivo estado del conocimiento científico poseía en su época. Según el biólogo francés Lamarck, que vivió antes de Darwin, las criaturas vivas transmitían los rasgos que adquirían durante su vida a la siguiente generación, y estos rasgos acumulándose de una generación a otra provocaban la aparición de nuevas especies. Por ejemplo, según Lamarck, las jirafas evolucionaron de los antílopes; esforzándose por comer hojas de árboles altos sus cuellos fueron estirándose de una generación a otra.
Darwin da también ejemplos similares en su libro “El origen de las especies”. Por ejemplo, dice que algunos osos que se introducían en el agua para buscar comida se transformaron en ballenas con el paso del tiempo (51).
Pero no obstante, las leyes de la herencia descubiertas por Mendel y verificadas por la genética que floreció en el siglo XX, finalmente destruyeron la leyenda de que los rasgos adquiridos se trasmitían a las generaciones subsiguientes. De esta forma la selección natural perdió sustento como mecanismo evolutivo.
EL NEODARWINISMO Y LAS MUTACIONES
Para encontrar una solución, los darwinistas propusieron a finales de la década de 1930 la “moderna teoría sintética” o, como se la conoce más comúnmente, el neodarwinismo.
El neodarwinismo agregó las mutaciones, que son distorsiones producidas en los genes de los seres vivientes como resultado de factores externos tales como radiaciones o fallas en la replicación, como “causa de las variaciones favorables” además de la selección natural.
Actualmente el modelo evolucionista que persiste en el mundo es el neodarwinista. La teoría sostiene que los millones de seres vivientes presentes en la tierra son el resultado de mutaciones o desórdenes genéticos por medio de los cuales se fueron provocando cambios en numerosos órganos complejos de estos organismos, tales como oídos, ojos, extremidades, alas, etc. Sin embargo, existe un hecho científico innegable que socava esta teoría: las mutaciones no provocan una evolución en los seres vivientes; por el contrario siempre los perjudican.
La razón para esto es muy simple: el ADN tiene una estructura muy compleja y las mutaciones azarosas sólo pueden dañarla. El genetista americano B. G. Ranganathan explica esto como sigue:
“Las mutaciones son pequeñas, azarosas y dañinas. Ocurren raramente y lo mejor que puede pasar es que sea ineficaz. Estas cuatro características de las mutaciones implican que no pueden conducir a un desarrollo evolutivo. Un cambio al azar en un organismo altamente especializado o es dañino o es fútil. Un cambio al azar en un reloj pulsera no puede mejorarlo. Lo más probable es que lo perjudique o, en el mejor de los casos, que sea insustancial. Un terremoto no mejora una ciudad, la destruye” (52).
No es sorprendente que ningún ejemplo útil de mutación, esto es, que se haya constatado que mejoró el código genético, se haya observado hasta ahora. Todas las mutaciones han probado ser nocivas. La mutación, que se presenta como un “mecanismo evolutivo”, es realmente un incidente genético que daña a los seres vivientes y los incapacita. (El efecto más común de mutación en los seres humanos es el cáncer). No caben dudas que un mecanismo destructivo no puede ser un “mecanismo evolutivo”. La selección natural, por otro lado, “no puede hacer nada por sí misma” como también Darwin aceptó. Esto nos indica que no hay “mecanismos evolutivos” en la naturaleza, y si no existen difícilmente alguien pueda imaginar cómo el proceso llamado evolución tendría lugar.
Etpoursimouve, parece que hemos llegado a un entendimiento, y, ya ve usted, ¡a base de argumentos! Alguno habrá que se quede con la boca abierta...
Mi incomodidad con el tema venía con el titular "Los ancestros del hombre comenzaron a caminar primero en los árboles", que me parece que tiene que ser, al menos, una mala interpretación periodística de los resultados de una investigación que, por otro lado, no dudo que sea rigurosa y coherente.
El problema del bipedismo es, efectivamente, aún más interesante si tomamos en cuenta su probable aparición en un ambiente boscoso. Inspira cuestiones sobre qué tipo de bosque sería, sobre cómo sería la morfología del protohomínido en cuestión, qué uso haría de su ambiente, cuál sería la secuencia de los cambios morfológicos, etc, etc. Más aún teniendo en cuenta que, según me parece, no hay referentes actuales (en forma de animales vivos) que nos puedan guiar con su ejemplo (el orangután es pesado y arborícola, pero no bípedo).
Como dices, hay que dejar trabajar a los profesionales y esperar los resultados con la mente abierta, pero también con sentido crítico.
Otro codial saludo.
Apomios, estás exponiendo críticas a la teoría científica de la evolución provenientes, precisamente, del campo científico. Yo esas críticas ya me las conozco, así como también los esfuerzos que los propios investigadores están haciendo para resolver las cuestiones que tienen abiertas.
Lo que me interesaría es saber otras cosas, como, por ejemplo, qué dijo concretamente el profeta sobre el ADN, el signo de las mutaciones, las leyes de la herencia, los tres tipos de selección natural, y, ya de paso, sobre la cura del cáncer... A ver si va a resultar que las respuestas ya están escritas en el Corán y los pobres científicos están perdiendo el tiempo con sus libracos y sus probetas.
Que bonito que todos tiramos la máscara.
Del trono de gigantes a la Hispania prerromana monolingüe y con todo quisqui pagando en moneda, faltaría más.
Por cierto, ateo y a mucha honra. Para dar fe de algo no hace falta ser creyente. Solo haber pasado unos años en una facultad de Derecho, y alguna otra cosa más.
A todo esto, si A es la nueva encarnación de otro druida, y si Z (quien a su vez es probablemente encarnación de I.), ya ha aparecido en danza ¿cuánto tardará en aparecer S? Solo falta JM para que toda la tropa esté al completo.
Apomios:
Acerca de que las mutaciones no producen ventajas adaptativas, espero que nunca tengas una infección resistente a un antibiótico... o acaso una mutación que permite a una bacteria X resistir un antibiótico ¿No es un claro signo de ventaja adaptativa?
La reorganización al azar de los genes de los anticuerpos humanos nos proporcionan los anticuerpos necesarios para eliminar muchísimas infecciones, otro claro ejemplo de como alteraciones en el DNA resultan en una ventaja adaptativa.
Otro ejemplo lo tienes en Pseudomonas, las cuales "son capaces" de modificar regiones de sus genes para producir nuevas proteínas para "alimentarse" de nuevos substratos.
Acerca de este y otros argumentos comunes del creacionismo, acudan a un interesante artículo en scientific american:
"15 Answers to Creationist Nonsense
Opponents of evolution want to make a place for creationism by tearing down real science, but their arguments don't hold up"
By John Rennie
http://www.sciam.com/article.cfm?articleId=000D4FEC-7D5B-1D07-8E49809EC588EEDF&chanId=sa013&modsrc=most_popular
Información eliminada por el Administrador Silberius
Motivo: Mensaje repetido
Apomios05 de jun. 2007Klenow
Cuando se habla de mutaciones en este contexto, hace referencia a que un lobo siempre tendra que ser un buen lobo para sobrevivir, no convertirse en un inocente perro faldero por arte de magia.
Apomios05 de jun. 2007¿Que necesidad tiene un lobo, en convertirse en un inocente chiwawa, eso es evolucion?
No dice la teroria de la evolucion, que los perros actuales evolucionaron del lobo, no seria eso una involucion?
Un lobo tendra que ser el mejor lobo para sobrevivir, gracias a la seleccion natural y el gen de la herencia.
Que yo sepa, ahora mismo hay en el mundo muchos más perros que lobos.
Apomios05 de jun. 2007Pero no por seleccion natural, sino por seleccion artificial querido amigo.
mira tío! a ti la mente es que no te da, no llegas al umbral mínimo para comprender entender e interpretar ciertas cosas, el lobo es un animal con una plasticidad social de la ostia!,eso le permitio establecer una relación con el hombre primitivo insospechada hasta entonces, y esta relación además adquirió tintes de interdependencia. A nivel morfológico los cambios que se produjeron fueron inducidos por el hombre ya que la selección natural dejo de actuar sobre el lobo para actuar sobre el una nueva selección ( la artificial) en el lobo, la presión selectiva ejercida por el hombre origina nuevas tendencias de lo que iba buscando tanto a nivel morfológico como a nivel de comportamiento. Pero esto que no comprende se hace a través de la variabilidad, ya que no existe 2 individuos genéticamente iguales (salvo las excepciones, como los univitelinos), y que por muy iguales que veas dos lobos, hay diferencias!
Ahora vamos a analizarlo no a nivel morfológico, sino a nivel de estrategia, para un animal especializarse es sinónimo de garantía de supervivencia si el medio de especialización es continuo e invariable (cosa que nunca pasa), cuando el lobo se asoció al hombre se produjo una especialización, el agente especializador es este caso fue el hombre, el cual proporcionaba los recursos que el lobo precisaba (basicamente comida, reducción de la mortalidad, mas eficacia reproductiva, etc, etc, etc) de esta manera el cociente de beneficios le salia positivo por lo que la asociación fructifico y la especie se dejo moldear por su especializador, que en este caso infería en el bajo criterios artificiales.
Esto para que te sirva de ejemplo es lo mismo cuando ves una especie especialista como por ejemplo el Guepardo, altamente especializado en uno o dos tipos de presas (impalas y/o gacelas thompson), y ves que su cuerpo es el resultado de una carrera armamentista entre el guepardo y la gacela tompson,pero que pasa si el especializador se extingue?, pues que muy probablemente el especialista también! por si no lo sabes esto se demuestra matematicamente con ecuaciones logísticas definiendo las variables K=especialista , r = generalista, así que mejor ahorrate la discusión y la replica porque esto si que es tan demostrable como 2 y 2 son 4, y además dudo que sepas interpretarlo,
¿Y cuál es la diferencia?
Lobos más bien cutrecillos, tirando a mansos, que en la manada no hubieran prosperado, se encuentran en un medio ambiente en el que hay una especie de primate que les permite sobrevivir, y, posteriormente, prosperar, hasta diferenciarse en una especie diferente: de Canis lupus a Canis familiaris.
Se me ha adelantado Etpoursimuove.
Apomios05 de jun. 2007Y como puede convertirse un lobo de 50 kg en un york shire de 100 gramos de peso, con una morfologia completamente diferente?
Apomios05 de jun. 2007etpoursimouve
Hoy, a las 13:15
A nivel morfológico los cambios que se produjeron fueron inducidos por el hombre ya que la selección natural dejo de actuar sobre el lobo para actuar sobre el una nueva selección ( la artificial) en el lobo, la presión selectiva ejercida por el hombre origina nuevas tendencias de lo que iba buscando tanto a nivel morfológico como a nivel de comportamiento.
¿Entonces las razas de perros como dices no son evolucion y son una seleccion artificial?
¿Si encuentran un perro fosilizado en el futuro, diran que es evolucion o seleccion artificial?
Apomios05 de jun. 2007Te dire algo, conozco muchos pastores que intentaron trabajar con cachorros de lobo para que hicieran la funcion de perros pastores y poder enfrentarse a los lobos salvajes con las mismas armas. Te dire lo que sucedio, hasta la fecha ninun cachorro de lobo educado con el hombre a podido cumplir la faena de un perro pastor, terminaban comiendose el ganado.
Es evidente que la teoría de la evolución hace aguas por todas partes.
* Premisa 1: el Monstruo de Espagueti Volador es un ser que tiene toda perfección.
* Premisa 2: la existencia es una perfección.
* Conclusión: por lo tanto el Monstruo de Espagueti Volador existe.
* Premisa 1: la existencia de todo lo que comienza a existir tiene una causa.
* Premisa 2: el universo comenzó a existir.
* Conclusión 1: por lo tanto la existencia del universo tiene una causa.
* Premisa 4: como no hay una explicación científica que pueda elucidar la causa del origen del universo, esta causa debe ser sobrenatural, o sea el universo fue creado por un dios.
* Conclusión 2: por lo tanto, un dios existe (o no).
* Premisa 5: los dioses siempre han creado a los humanos a su propia imagen y semejanza.
* Premisa 6: el cerebro de los humanos parece una fuente de espaguetis.
* Conclusión 3: por lo tanto, el Monstruo de Espagueti Volador es el único Dios verdadero.
Todo esto es completamente irrefutable. Así que malignos científicos ya os podeis ir marchando a vuestros malignos laboratorios fasciocomunistas.
Apomios05 de jun. 2007Y te dire el pq, se llama gen de la herencia.
Apomios05 de jun. 2007Asi es Protion9, lo que no entiendo es como se creen todo ese cuento sin pensar por si mismos, creo que estamos en la epoca de creer por lo que te dicen y no por lo que se deduce por logica.
Apomios05 de jun. 2007Mas que malignos, diria inocentes fanaticos que se lo creen todo
Copio y pego un extracto de un trabajo de Ecologia
Dinamica y efectos de la predación
2.1 Efectos:
Estos efectos suelen ser perjudiciales en la poblacion de presas, pero existen casos que si son perjudiciales para la presa no lo son tanto para el conjunto de la poblacion, esto se debe a que si la densidad de la poblacion de presas es lo suficientemente alta como para que se esten dando fenomenos de competencia intraespecifica, el efecto de la depredacion hara efectiva una reduccion del numero de individuos que compiten por el alimento o el espacio. Por esto la competencia intraespecifica se vera aliviada.
De cualquier forma los depredadores van a determinar en ultima instancia la abundancia de las presas, o viceversa. Para ello existe un modelo matematico que intenta explicar como funcionan este tipo de interacciones llamado Lotka – Volterra
2.2 Modelo de Lotka y Volterra:
Recibe su nombre de los que fueron sus creadores, es un modelo que posee dos componentes; C, el numero de individuos existentes en una poblacion de consumidores, y N, que es el numero de individuos existentes en la poblacion de presas.
De este modo se puede decir que en ausencia de consumidores las presas aumentaran su poblacion exponencialmente:
dN∕dt=rN
sin embargo los individuos presa seran eliminados con una frecuencia que dependera de los encuentros que se den entre la presa y su consumidor, esta frecuencia aumentara en funcion del aumento de depredadores C, yo presas N.
Los encuentros fuctiferos y la tasa de consumo seran:
dN∕dt=rN-aCN
Donde aCN es la tasa de consumo. Este modelo tambien explica que en ausencia de alimento los depredadores disminuiran su eficacia y moriran de hambre, por lo que su numero decrecera exponencialmente:
dC∕dt=-qC
Donde q es la tasa de mortalidad. Pero esto se ve contrarrestado por el nacimiento de nuevos depredadores, cuya tasa depende de:
i) La tasa de consumo de alimento
ii) Eficacia, f, para convertir el alimento en descendientes
Por lo que la tasa de natalidad es faCN
dC∕dt=faCN-qC
Este modelo resulta muy util para señalar las tendencias en las relaciones depredador presa, de forma que se estudian las oscilaciones entre ambos linajes. Este mecanismo de oscilaciones acopladas posee una serie de retrasos en el tiempo, esto se debe a que los periodos de abundancia de presas no dan tiempo a satisfacer a la poblacion de presas. De este modo se produce un retraso en el tiempo de pocas presas y pocos depredadores y asi sucesivamente.
3. Preferencias alimentincias
3.1 Preferencias y tipos de consumidores:
Existen diferentes modelos de comportamiento en los consumidores ala hora de elegir o seleccionar aquello que va a ser consumido.
En lineas generales vamos a encontrar 3 tipos de consumidores. Monofagos (solo consumen un tipo de especie), olifagos ( consumen solo varias especies), polifago (consumen un rango muy alto de especies). Es decir, las especies monofagas seria las tambien llamadas especialistas, las polifagas son las generalistas y las olifagas serian las que tienen un rango intermedio.
Sin embargo las especies denominadas “generalistas” no toman cualquier cosa que se encuentren como alimento. Sino que aceptan solo una estrecha gama de los recursos existentes ( un raton es un recurso pero un leon no se lo come). Es decir, dentro de la generalidad de una especie polifaga u olifaga, existe una preferencia a la hora de elegir alimento.
Estas preferencias estan siempre sujetas a lo provechosas que puedan llegar a resultar para el individuo que las consumen, dicho de otro modo: Los individuos consumidores tenderan a escoger las presas mas energeticas.
Pero esto no siempre resulta asi, en la alimentacion de los consumidores herbivoros muchas veces la eleccion del alimento esta condicionada la concentracion de las toxinas que las plantas pueda contener y esto puede llevar en ambientes donde las plantas estan bien equipadas a la herbivoria a una dieta mixta. En estos casos una dieta mixta mantiene las concentraciones de toxinas a niveles adecuados, ya que cada especie vegetal sintetiza una toxina propia.
3.2 Amplitud de la dieta:
El primer trabajo que desarrolla un modelo en la amplitud de la dieta fue el llamado teoria del aprovisionamiento optimo (Mc Arthur & Pianka). En esta teoria se argumenta que para obtener energia todo depredador necesita invertir tiempo y energia, primero en buscarla y luego en manipularla, por ello los investigadores consideraron que la amplitud de la dieta dependia en ultima instancia de la capacidad de manipulacion, de esta forma cuando un depredador generalista busca presas encuentra un alto numero, mientras que un especialista solo consume la presa que prefiere. Esto plantea un gran peoblema si el individuo es un especialista, dado que solo consumira presas provechosas, los tiempos de busqueda pueden resultar altos y puede invertir mucho tiempo y energia. Mientras, si es un generalista los tiempos de busqueda seran breves.
Segun esto la teoria hace 4 predicciones:
i) Los buscadores deberian ser generalistas
ii) Los manipuladores especialistas
iii) La especializacion deberia ser mayor en ambientes productivos
iv) La abundancia de los tipos de presas no provechosas es irrelevante
La primera de estas afirmaciones parece verse confirmada si tenemos en cuenta a las especies de aves paseriformes insectivoras como la curruca cabecinegra Sylvia melanocefala, cuya dieta esta compuesta esencialmente por diminutas larvas de insetos, esto requiere un tiempo muy corto de manipulacion. Por lo que una dieta amplia le conlleva mas ventajas que desventajas. Por el contrario depredadores como el leon africano Phantera leo, viven casi la totalidad de su vida a la vista de sus presas,
por lo que el gasto en encontrarlas es casi nulo e invierten mas en la manipulacion, la persecucion puede llegar a ser mas larga que la eleccion de la presa y por tanto se especializan en las presas provechosas (enfermas, jovenes y viejos).
3.3 Carrera de armamentos:
De cualquier forma la polifagia parece ofrecer mayores ventajas. La inversion en busqueda es menor, morir de hambre es mas dificil y aun asi se dan en la naturaleza casos en los que asociaciones depredador-presa lejos de ampliar las dietas se han visto reducidas a la minima expresion, es decir una planta o presa es consumida por un solo tipo de herbivoro o depredador, y a su ven estos solo pueden consumirlos a ellos. Esto solo se entiende en el contexto de la evolucion y en este caso las parejas depredador presa no solo han evolucionado si no que han coevolucionado. La idea es que se produce una “carrera de armamentos” evolutiva en la que existe una mejora progresiva en la efectividad de caza de un predador para con una presa cualquiera pero a su vez esta tambien va mejorando su capacidad de escape. Esta idea nos sugiere que si trasladasemos un predador de ratones del Eoceno quizas se moriria de hambre intentando capturar a uno actual, y ademas nos explica porque ciertos insectos se parecen a hojas o la semejanza entre una polilla y una corteza de arbol, y esto es probable gracias a una larga carrera de armamentos entre insectos y aves insectivoras dispuestas a discriminar entre las verdaderas presas y sus modelos no comestibles.
3.3.1 Factores que otorgan ventajas:
A veces se dan situaciones donde algunas de la partes implicadas juega con ventajas. Sin que con ello se quiera decir que esta parte de la contienda vaya a ganar la carrera de armamentos, lo que ocurre es que una de las partes experimenta procesos de evolucion mas altos que la otra. Cada vez que uno de los dos linajes experimente una adaptacion, disfrutara de un periodo de relajacion ya que el otro linaje disminuira su eficacia durante algun tiempo. Existen varios factores que otorgan ventajas a una de las partes:
i) Las presiones selectivas son desiguales. Para una presa y un depredador las presiones selectivas son desiguales ya que uno corre por su vida mientras que el otro lo hace por su cena. Esto significa que el conejo que no escape a su perseguidor jamas se reproducira, mientras que el zorro si lo hara aunque equel dia no llegase a capturar el conejo. Por ello la asignacion del conejo a “escapar del zorro” sera mayor que la del zorro “atrapar conejo”, por lo que la eficacia global es mayor en las presas que en los depredadores y parten siempre con una ligera ventaja.
ii) Efecto enemigo raro. Aqui uno de los dos linajes explota al otro limpiamente, y lo hace con el efecto del engaño. En las luciernagas, los machos y las hembras se reunen para la reproduccion. Algunas luciernagas del genero Photuris imitan las señales de las hembras del genero Photinus de manera que cuando los machos se acercan para un posible cortejo, estos son capturados y devorados. Pero podriamos preguntarnos por que estas se dejan engañar por las “femme fatale”, la razon es simple, el depredador es una especie mas rara que la presa.
iii) Presupuestos adaptativos. Cualquier adaptacion es costosa. Si un linaje se centra en una carrera de armamentos concreta obviara otros asuntos, un Guepardo puede evolucionar hacia una carrera aun mas rapida pero obviara quizas la maniobrabilidad y la resistancia para arrastrarla y consumirla.
iv) Tasas adaptativas. Segun el registro fosil, los diferentes grupos taxonomicos no han evolucionado a la misma velocidad. Una salamandra actual se parece mas a un anfibio de Carbonifero que un ave actual a un Archaeopteryx, y ya sabemos que estas dos ultimas estan mas cerca en el tiempo. Los factores que pueden acelerar estos procesos son muy diversos; varianza genetica , aislamiento poblacional, tiempo de generacion cortos, etc, etc.
4. Parasitismo, parasitos y parasitoides
4.1 Clasificacion y definicion:
Como ya se definio en el apartado 1.3, los parasitos son organismos que consumen solo una parte de los recursos de un organimo, suelen causarles daño y a veces la muerte. Sin embargo su clasificacion no ha sido tan sencilla como su definicion, asi que tomaremos la diferenciacion entre micro y macroparasitos, de forma general:
i) Los microparasitos son pequeños, muy numerosos, y se multiplican en el interior de sus huespedes. La mayoria de estos son intracelulares y son los responsables de las infecciones bacterianas y viricas. Gripe, malaria, y sarampion son algunos ejemplos.
ii) Los macroparasitos son bastante diferentes a los anteriores, generalmente poseen una fase de crecimiento en el interior del huesped, y otra de multiplicacion en el exterior, donde la descendencia busca nuevos hospedadores que parasitar. Piojos, pulgas, garrapatas, duelas, y tenias son tambien algunos ejemplos.
4.2 El parasito y su habitat:
Lo que hace a un parasito un parasito, es que su habitat es un organismo vivo. Esto hace que su habitat este en continua lucha contra el organimo parasitante, de hecho combatimos (en el caso de los mamiferos y aves) algunos microparasitos elevando la temperatura corporal, desarrollando reacciones inmunitarias, digiriendolo, etc, etc.
Considerando pues a los huespedes como habitas Janzen (1968,1973) introdujo la analogia de pensar en los huespedes como islas colonizadas por parasitos. De esta forma un individuo parasitado es en ultima instancia una isla colonizada, esto lleva al estudio del parasitismo desde el punto de vista de la ecologia, al mismo concepto que el estudio de la biogeografia de islas.
4.2.1 dispersion y colonizacion:
Las diferentes especies de parasitos tienen modelos de dispersion diferentes. En el caso de los microparasitos el contagio (“colonizacion”) se produce de manera proporcional al numero de encuentro que puede darse en una poblacion de posibles huespedes, por ello la propagacion de una epidemia esta relacionada con la distancia que exista entre huespedes.
El exito de la colonizacion tambien esta relacionado con la alta especificidad que suelen presentar los parasitos, ya sabemos que los organismos se encuentran muy restringidos debido a su especializacion a determinados ambientes. Como ya hemos visto en el anterior capitulo, los depredadores estan especializados en sus presas y aun siendo generalistas o polifagos la gama de presas para las que esta diseñado un predador reduce mucho el rango de estas. En los parasitos resulta si cabe aun mas intensa la relacion con su huesped, y normalmente es facil encontrar parasitos que estan restringidos a una sola especie de huespedes, y esto es asi porque el parasito esta dentro de una estrategia evolutiva estable (recordemos el apartado de la carrera de armamentos).
Por otro lado los parasitos no solo se especializan en una especie o genero, sino que ademas lo hacen sobre algun tejido especifico. Esto nos lleva a dar una nueva clasificacion:
i) Los Ectoparasitos toman su alimento desde el exterior, pulgas y garrapatas en animales y afidos en las plantas;
ii) Un grupo intermedio que desarrollaria su actividad en la cavidades internas, como son los nematodos que desarrollan su actividad en los canales alimenticios.
iii) Y por ultimo los endoparasitos, como la malaria, que desarrollan su actividad en el interior de los tejidos.
4.3 Adaptaciones al parasitismo:
La estructura general de los endoparasitos refleja normalmente las condiciones menos rigurosas, limitadas y uniformes del ambiente en el que viven. Los sistemas locomotores suelen estar reducidos, y los organos sensitivos que se encontrarian en organismos de vida libre, a veces estan reducidos o incluso han desaparecido. Y el sistema nervioso tambien se reduce a su vez.
Sin embargo los parasitos han desarrollado varias especializaciones para protegerse del hospedador. Algunos se han especializado en zonas donde el sistema inmune no llega, como es el tubo digestivo, otros poseen fases de desarrollo enquistadas y otros incluso utilizan sistemas de mimetismo (como los insectos que se parecen a hojas) con el sistema inmune y adoptan los antigenos en su superficie para no se reconocidos por el sistema inmunitario, este es el caso de los trematodos.
Un parasito con exito no mata a su hospedador, o al menos no deberia hacerlo, porque es obvio que la muerte del hospedador acarrea la suya propia. Si lo hace tendra que tener tiempo para que escape el o sus huevos.
4.3.1 Evolucion del parasitismo:
Los ectoparasitos han evolucionado a partir de especies que inicialmente eran comensales u ocupantes ocasionales de la superficie de los hospedadores. Algunos parasitos han evolucionado a partir de antecesores que estaban preadaptados a una existencia parasita. Por ejemplo, algunas moscas parasitas ponen sus huevos perforando la piel del ganado, y estas estan estrechamente relacionadas con especies que ponen sus huevos en la carne de animales muertos.
Por otra parte la boca de los hospedadores juega un importante papel en la evolucion de algunos endoparasitos. Los antecesores fueron ingeridos accidentalmente y sobrevivieron. La supervivencia podria haber originado una seleccion rapida de las carcteristicas que mejoraron las relaciones parasitarias. En aquellos que poseen hospedadores intermediarios, estos fueron quizas probablemente los hospedadores primitivos. Cuando el primer hospedador fue comido por el segundo, el parasito volvio a sobrevivir pero las adaptaciones que hicieron posible su supervivencia en el primero tuvieron que ser conservada en su ciclo biologico.
4.4 Efectos sobre la supervivencia, el crecimiento y la fecundidad.:
Una vez que el parasito o los parasitos han infestado un individuo este reducira su tasa de crecimiento y de fecundidad, asi como aumentara su grado de depredacion, de hecho tambien puede llegar disminuir la eficacia competitiva del hospededor frente a otros competidores. Un importante hecho es que puede debilitar a un competidor agresivo, y favorecer a otro no agresivo que no este infectado.
Parece que el principio “causar daño al huesped” es condicion indispensable para la definicion de parasitismo. Pero esto no siempre es tan claro y a veces el daño que se produce es mas sutil de lo que cabria esperar. Por ejemplo en la especie papamoscas cerrojillo (Ficedula hypoleuca) puede ser infectado por un Trypanosoma sanguineo, este reduce ligeramente la longitud de su cola (que es sinonimo de atrayente sexual para la hembra), de modo que cuado quieren aparearse son menos virtusos para las hembras. Pero existe un experimento en una lagartija australiana (Tiliqua rugosa), cuya longevidad no se hallaba correlacionada con la carga de garrapatas que arrastraban, y los individuos que se apareaban presentaban mayor numero de ectoparasitos que los que no se apareaban.
En general los organismos que son resistentes al parasitismo evitan los costes que produce ser parasitado, de la misma forma que sucede con la resistencia con enemigos naturales. Es decir en ausencia de parasitos una especie A susceptible al parasito sera mas competitiva que una especie B resistente al parasito. Los organismos estan sujetos a los costes de susceptibilidad y resistencia de igual forma que en el apartado 3.3.1-iii, cuando citaba el ejemplo del Guepardo.
4.5 Un caso especial, parasitismo de cria:
Hasta ahora hemos visto relaciones parasitarias donde el recurso que se explota es el huesped en si mismo robandole sus tejidos para la nutricion del parasito o aprovechando su maquinaria celular yo energia, y en casi todos los casos las relaciones filogeneticas entre ambos linajes solian estar muy alejadas, bacterias-mamiferos, artropodo-reptil, etc, etc. Pero en el caso del parasitismo de cria, las relaciones filogeneticas suelen estar muy proximas e incluso a veces se dan en el seno de la misma especie.
Se conocen ejemplos en hormigas que capturan larvas de otras especies a las que luego someten como obreras, pero son las aves las que proporcionan los ejemplos mas conocidos. Muchas hembras de gorrion domestico (Passer domesticus) son incapaces de aparearse con un macho que posea un territorio, por lo que se aparean con varios machos dominantes y depositan sus huevos en los nidos de otras hembras (parasitismo intraespecifico), con ello evitan los costos de la reproduccion delegandolos a otras parejas. Este tipo de parasitismo puede llegar a ser ocasional en numerosas especies de aves (tambien se han descrito casos en los que algunas Anatidas realizan esta actividad), pero en ciertos grupos ha pasado de ser facultativo a ser un hecho obligado. Tal es el caso que en el cuco europeo (Cuculus canorus) depende para su reproduccion de otras especies por completo (parasitismo interespecifico). Normalmente se deposita un unico huevo y en la mayoria de los casos el polluelo intenta monopolizar los cuidados de los padres adoptivos, matando (arrojando del nido) a sus “hemanastros”.
Ahora bien, este modelo de parasitismo se aproxima bastante a los modelos de parasitismo mas conveccionales, y tambien se dan procesos de carreras armamentisticas. La presencia del parasito disminuye la eficacia del hospedador, por lo que este es seleccionado para detectar su presencia y rechazarlo: bien expulsandolo del nido o bien negarse a cuidar de el. El parasito por su parte desarrolla adaptaciones para engañarlo y producir huevos y pollos mimeticos, el parasito juega con ventaja. La presion selectiva (apartado 3.3.1-i) es mas alta para el parasito ya que este se juega la vida mientras el hospedador no, si bien la de su descendencia si. Una cria de cuco puede dedicar gran parte de sus recursos hacia la adaptacion y engañar a sus padres falsos yo excluir a sus hermanastros . Sin embargo, el hospedador debe dedicar sus adaptaciones no solo ha criar mal un cuco sino a criar bien a sus hijos propios, con lo cual ambos linajes corren carreras de armamentos diferentes. Aqui suelen ganar los parasitos y estos parten con ventaja.
5. Simbiosis y mutualismo
5.1 Definicion:
La simbiosis es un termino que significa “vida en comun”, esta definicion se creo para agrupar bajo un mismo nombre a todas aquellas especies que mantienen una estrecha relacion con otros organismos de diferentes especies. A diferencia del parasitismo, las relaciones que aqui se establecen suelen ser beneficiosas para ambos actores. Pero muchos organismos se benefician unos de otros y muy discutidamente puede decirse que lleven una vida en comun (clasico ejemplo del insecto buscando polen en la flor.Mutualismo). Otras veces el beneficio va solo en una direccion aunque sin perjuicio del otro (comensalismo)
5.1.1 Mutualismo:
Se trata de una asociacion de un par de especies (generalmente), y cuyo encuentro acarrea siempre un beneficio mutuo. El mutualismo no implica una relacion fisica entre la pareja, por lo que los mutualistas no son necesariamente simbiontes. Por ejemplo; el clasico ejemplo de la abeja (Apis melifera) y la margarita. Es indudable que ambos se ven beneficiados del encuentro, la abeja obtiene alimento y la flor a cambio del polen cedido es fecundada. Sin embargo estas interacciones mutualistas no son consideradas como simbioticas. La frontera entre mutualismo y simbiosis esta poco clara pero de modo general se puede establecer que el mutualismo no implica una asociacion fisica-obligada y en la simbiosis esta asociacion se da de manera mas patente.
5.1.2 Algunos ejemplos de relaciones mutualistas:
i) El pez limpiador establece un territorio que podriamos denominar “estaciones de limpieza”. Los clientes (otros peces) los recocen y se acercan a estos puntos cuanto mayor es su grado de parasitismo. Con esta relacion mutualista los peces limpiadores obtienen alimento y el pez visitante se libra de sus parasitos.
ii) Otro ejmplo curioso es el del pajaro de la miel africano (Indicator indicator) y el tejon melifero (Melliovora capensis). Este pajaro es capaz de encontrar los escondidos panales de las abejas pero por si solo es incapaz de explotarlo como recurso, de forma que cuando localiza uno se concentra en la busqueda de un tejon y lo atrae hasta el panal. El mamifero rompe el panal y nuestro astuto pajaro se alimenta de las sobras.
iii) Los langostinos del genero Alpheus excaban madrigueras y el pez gobio Cryptocentrus se aprovecha de ellas en un ambiente donde las cavidades para la proteccion son ausentes.El langostino que es ciego mantiene contacto con el pez atraves de sus antenas y este le avisa en caso de peligro. Las ventajas que ambos obtienen son claras.
iv) Los mutalismos de dispersion de semillas son el resultado de una larga coevolucion entre plantas aves y mamiferos. Los beneficios de que la planta obtiene son unicamente la dispersion de semillas, pero no es poco ya que con ello las plantas se alejan aleatoriamente de su area “materna” y colonizan nuevos territorios, por otro lado los animales que la consumen obtienen alimentos ricos en azucares y lipidos. La costosa inversion que tiene que hacer las plantas esta en el fruto, que se ve recompensado por la dispersion de las semillas.
5.1.3 Mutualismos que implican cuidado de ganado y cultivos:
Las hormigas son un grupo especializado en este tipo de mutualismos, en areas del Amazonas existen generos como Atta y Acromyrmex que se dedican a recolectar hojas de arboles y asi cultivar en ellas hongos del genero Hypomyces, el hongo es cultivado para el consumo pero con la hormiga su supervivencia esta asegurada. Otras hormigas mas conocidas como las de jardin (genero Lasius) establecen una actividad mutualista con insectos hemipteros. Los pulgones parasitan las yemas de las plantas, y las hormigas como ya sabemos precisan de un aporte de azucares en su dieta ( que dicho sea de paso es simpre omnivora), precisamente estos “animalillos” pueden proporcionarles toda esa “azucar” que necesitan. Los pulgones chupan mas de lo que necesitan puesto que lo que ellos buscan realmente es nitrogeno, el subproducto es expulsado por la parte posterior superior del abdomen y las hormigas solo tienen que recogerlo. Este relacion otorga a las hormigas “comida” y a los pulgones una estimable defensa contra cualquier prededor.
5.1.4 Otros tipos de mutualismos:
Como podemos ver existen relaciones mutualistas a la carta, intentar por tanto sintetizar todos en un trabajo como este seria tarea imposible y cuanto menos redundante. Por ello y al margen de los que merecen la pena ser mencionados (como el rumen ), citare algunos de las relaciones mutualista de mayor importancia:
El rumen: La mayoria de los intestinos de los animales es un microcosmos de vida bacteriana y protozoos. En ellos se da unas condiciones de Ph y temperatura mas o menos constantes y se suelen dar condiciones de anaerobiosis. Los mamiferos pequeños tienen asignado una parte de sus intestino para la fermentacion (el ciego) de los vegetales que comen y en animales grandes como el caballo el ciego es el lugar mas importante para la fermentacion.
En los animales herbivoros rumiantes (ciervos, vacuno, antilopes, etc.) la camara fermentativa se encuentra a nivel del estomago, esta dividido en 4 compertimentos. La primera masticacion conduce el alimento a la camara del rumen de ahi pasa al reticulo a medida que su tamaño va disminuyendo, de ahi pasa a la tercera camara, el omaso, una vez ahi el animal regurguita y se produce una segunda masticacion que reduce aun mas las particulas vegetales. Lo verdaderamente curioso es la actividad microbiana que se da en el rumen, y los productos de la fermentacion que ellos generan es lo que alimenta al huesped. A su vez es el huesped el que genera las condiciones necesarias para este proceso. Estas bacterias y protozoos son anaerobias estrictas y si entran en contacto con el oxigeno mueren. La mayoria de las especies que forman el rumen poseen tal especializacion que no podrian vivir fuera de el.
La cuestion de esta relacion es la carencia por parte del rumiante del aparato enzimatico para degradar la celulosa y otras fibras indigeribles en la dieta de los rumiantes. Bacteroides succinogenes digiere solo celulosa, Ruminococcus celobiosa y celulosa, otros pueden digerir tambien el almidon como es el caso de Clostridium locheadii. Ahora bien, como ya dije antes el tracto digestivo de los animales puede ser un microcosmos de vida, por tanto la flora del rumen esta sujeta a fenomenos de competencia en una escala inferior, al tratarse ademas de un ambiente constante (como ocurre en los tropicos), los fenomenos de competencia lejos de darse deben ser muy intensos.
Los liquenes: Numerosos hongos han escapado a su habitual sistema de vida libre y se han asociado con algas. Estos hongos son los denominados hongos “liquenizados”, contienen una capa de celulas fotosinteticas cerca de la superficie (algas).
Los hongos obtienen del alga productos de la fotosintesis y en algunos casos nitrogeno. La ventaja para el alga no esta muy clara y aun es objeto de investigacion, solo en algunos generos como Trebouxia parece que debe existir alguna relacion de beneficio porque las formas de vida libre son raras.
No es asi en el caso del hongo, las formas liquenizadas son diferentes en morfologia de las formas libres por lo que el crecimiento de este se halla profundamente alterado por la presencia del alga, son ademas eficientes colonizadores y su dispersion como colonizantes es mayor cuando estan asociados.
5.2 Fijacion del nitrogeno y su relacion con el mutualismo:
Como ya sabemos el nitrogeno es fundamental para la vida y es el elemento constituyente de las proteinas, en la naturaleza lo encontramos formando parte de los seres vivos, de forma inorganica como amonio, nitrito y nitrato (formas asimilables por las plantas), y en la atmosfera donde esta en forma de N2. El nitrogeno atmosferico representa casi el 80% de todo el nitrogeno del planeta por lo que su disponibilidad para los seres vivos hace que sea uno de los elementos mas limitantes ( la mayoria de los organismos no pueden asimilarlo en su estado gaseoso). Sin embargo existen organismos capaces de asimilar el nitrogeno atmosferico.
Azotobacter, Azotococus, Rizobios, Clostridiun spp, Enterobacter y/o cianoficeas en general son algunos ejemplos de bacterias que sintetizan el nitrogeno del aire hacia formas asimilables por otros organismos (plantas). Pero la familia Rhizobiaceae es de todos los grupos la mas estudiada.
El contacto entre el rizobio y la planta huesped (siempre una leguminosa) se produce a nivel de los pelos radiculares de la raiz, la bacteria que se encuentra en estado de vida libre es estimulada para la multiplicacion por los exudados de la raiz. Estos exudados son los responsables de la activacion de una serie de genes de la bacteria, que controlan tambien el proceso de nodulacion. De forma general se desarrolla una colonia alrededor de un pelo radical este se riza y forma un tubo de infeccion por donde pasan las bacterias. Una vez estas han penetrado en las celulas del huesped, se produce la formacion del nodulo.
El huesped entonces desarrolla un sistema vascular “especial” que retira del nodulo los produtos de nitrogeno fijado y aporta nutrientes fotosinteticos. Dentro de los nodulos existe una proteina homologa a la hemoglobina, la leghemoglobina, que posee una alta afinidad por el oxigeno, favorciendo asi que el nodulo se mantenga anaerobico (hay que recordar que el equipo enzimatico de la fijacion de N2 se realiza en anaerobiois) para la nitrogenasa.
5.2.1 Efectos del mutualismo fijador de N2 atmosferico en los ecosistemas:
La fijacion de nitrogeno se reduce en los ambientes que son ricos en N2, pero si eventualmente excasea la fijacion se dispara. Esto conlleva que todas las especies vegetales con asociaciones mutualista obtengan un beneficio neto superior al resto de las plantas. Por lo que en los ambientes con deficiencia de nitrogeno, las leguminosas fijadoras llevan ventaja frente al resto de espescies. Sin embargo existe una paradoja en este hecho, y es que a medida que el ambiente es enriquecido con Nitrogeno, las otras especies vegetales tambien se benefician, por lo que la ventaja inicial se pierde ya que el ambiente ha sido mejorado para los competidores . Quizas esta sea una de las causas de porque las leguminosas (tanto herbaceas como arbustivas) no hayan dado lugar nunca a poblaciones dominantes en la naturaleza.
De cualquier forma la importancia de los organismos fijadores de nitrogenos (a parte de las relaciones mutualistas aqui descritas) es incalculable. Los aportes de nitrogeno que realizan de manera natural es la unica via de entrada de nitrogeno atmosferico al ecosistema.
5.3 Eucariotas, producto de una simbiosis:
Hemos podido ver hasta ahora un compendio de ejemplos que nos hace ver como organismos diferenten se hacen “socios” y en algunos casos pueden llegar ha convertirse en una especie de organismo fusionado. Pues aplicando este concepto Margulis (1975) desarrollo una atrevida hipotesis (hoy teoria endosimbiotica) sobre la evolucion de la celula eucariota.
Segun Margulis, la evolucion de los eucariotas debio producirse por interacciones de procaritas con otras procariotas, es decir quizas un dia una bacteria con el ciclo de Krebs fue incluida dentro de otra, sobreviviendo una dentro de la otra. Mas tarde se asociaron a otras que poseian cilios o flagelos y asi hasta que despego la evolucion de estos primeros procariotas. Esto explicaria el origen de los organulos celulares, cloroplastos y mitocondrias poseen ADN y gozan de cierta autonomia con respecto al resto de la celula.
6. Competencia Intraespecifica
6.1 Definicion, efectos y caracteristicas de la competancia intraespecifica:
Ya hemos visto que los organismos se comen, se engañan y se asocian, pero lo que aun no hemos visto es que tambien son victimas de sus propias actividades. No existe ningun organismo que viva aislado. Y al menos una vez en su vida tendra que reproducirse y siempre formara parte de una poblacion con los de su misma especie.
Las necesidades que un organismo posee son invariables con las del resto de individuos de su misma especie, esto unido a la necesidad de no perecer de inanicion o espacio para criar, puede acelerar la demanda de un determinado recurso y agotar la oferta del mismo. Entonces el recurso se vuelve raro y los organismos compiten por el.
Consideremos el siguiente ejemplo: una poblacion saltamontes que crece y vive en un campo de herbaceas. Para crecer y reproducirse los saltamontes necesitan 2 cosas materia y energia; pero para obtener esto tambien precisan energia para desplazarse. Si un saltamontes llega a una parcela donde no hay comida tendra que volar para encontrar otra parcela donde no halla llegado un saltamontes antes que el. Por ello la supervivencia del saltamontes estara condicionada a la capacidad de encontrar antes de que otro lo haga ya que en ello va su eficacia reproductora o su propia supervivencia.
Pero en realidad esta no es una competencia directa entre individuos, el saltamonte ni se percata de que hay o ha habido otros congeres en su parcela, solo responde al hecho de que no hay comida y la necesita. En otros ejemplos si puede darse una competencia directa entre individuos (Interferencia), el individuo que accede primero al recurso impedira su acceso a otros congeneres, Por ejemplo entre Septiembre y Octubre se registra en la peninsula Iberica el fenomeno de la berrea, los machos del ciervo (Cervus elaphus) luchan entre si para defender un haren de hembras y un territorio, aqui el recurso esta claro; hembras para la reproduccion, y este es un un recurso limitado.
De este modo la competencia por interferncia puede desarrollarse por un valor real (comida, espacio, hembras, etc) o por sustituto (la conquista de cierto territorio implica la posesion de un haren).
6.2 Natalidad y mortalidad:
De manera general se puede decir que la mortalidad de una poblacion en situacion de estres por una alta densidad, puede ser dependiente de la densidad, esto significa que la mortalidad se incrementa con la densidad. Esto es facil de entender si volvemos al ejemplo anterior del saltamontes. Si partimos de una poblacion X de saltamontes que crecen y se reproducen y controlamos su tasa de mortalidad, veremos que a medida que aumenta su densidad aumentara su mortalidad, y esto ocurre por hecho muy sencillo; a medida que crece la poblacion disminuye la tasa per capita de alimento para cada individuo, los fenomenos de competencia por el alimento aumentan, y solo sobreviven los individuos mas competitivos.
En el caso de la natalidad, los resultados obtenidos por los investigadores son casi una imagen especular de la mortalidad, por lo que los efectos de la competencia intraespecifica ocasiona de igual modo un descenso de la natalidad, y a largo plazo contribuye al mismo efecto sobre la poblacion que la mortalidad; la poblacion decrece.
Para este tipo de estudios sobre el crecimiento de poblaciones animales existen numerosos modelos matematicos que intentan explicar como evolucionan las poblaciones sujetas a fenomenos de competencia. Para poblaciones con estaciones de reproduccion que pueden escribirse mediante ecuaciones en “diferencias” (aumentan en pasos discretos), existen modelos que analizan como varia de tamaño la poblacion.
6.3 Ecuaciones en diferencias:
Estas ecuaciones nos sirven para teorizar como se comportara una poblacion bajo fenomenos de competencia intraespecifica. El modelo consta de dos ecuaciones:
t
Nt+1 = NtR y Nt= N0R
Nt representa el tamaño de la poblacion en el momento t, y R representa la tasa de reproduccion bajo la cual la poblacion crece.
Pero este modelo por si solo no nos resuelve nada, en el se describe una poblacion carente de competencia, por lo que si R$1, la poblacion creceria de manera exponencial ( esto es lo que ocurriria en la naturaleza con alimento y espacio “ad libitum”). Modificando ligeramente las ecuaciones y teniendo en cuenta los factores de competencia sobre la tasa de reproduccion, podemos obtener una situacion donde Nt sea un valor muy reducido (casi cero) y por tanto la competencia despreciable (recordar que los efectos de la competencia son patentes a altas densidades de poblacion), obteniendo:
Nt/ Nt+1= 1/R La tasa de reproduccion no queda altarada.
Pero a medida que Nt aumenta, la tasa de reproduccion se altera de manera que cuando el cociente Nt/ Nt+1 es igual a “0” la poblacion deja de crecer y solamente se sustituye en cada generacion. El tamaño de la poblacion por el que se produce este fenomeno es denominado capacidad de carga, K.
Si tenemos en cuenta la grafica A y B son los puntos que describen la tasa neta de reproduccion a medida que lo crece la poblacion. Nt/ Nt+1 se mide en el eje y, Nt en el eje x, y la interseccion es 1/R, la pendiente es por tanto
Nt/ Nt+1= 1-1/R/K x Nt + 1/R
Reorganizando:
Nt+1= Nt R/1+(R-1) Nt/K simplificando, (R-1) con una “a”:
Nt+1= Nt R/(1+aNt)
Este es el modelo definitivo, donde R a pasado de ser una constante a ser la tasa real de reproduccion, R/(1+aNt), que disminuye a medida que aumenta el tamaño de la poblacion Nt.
6.4 La ecuacion logistica:
Ya hemos visto muy a grosso modo el modelo matematico con ecuaciones continuas, y como este nos sirve para hacer estudios sobre el comportamiento de las poblaciones. Sin embargo para poblaciones donde las muertes y los nacimientos se producen de manera continuada, se utilizan modelos donde las ecuaciones mas apropiadas son las “logisticas”.
Aqui el crecimiento de una poblacion se denotara como dN/dt. Esto nos representa la velocidad de crecimiento N, en un tiempo t. Asi pues la tasa de incremento per capita es dN/dt(1/N), pero como esta expresion no recoge la competencia, suponemos que lo que medimos es en ausencia de competencia:
dN/dt=rN
En esta expresion donde no existte competencia, la poblacion crecera exponencialmente como se ve en la grafica. Para ajustar el modelo a la realidad debemos introducir la competencia intraespecifica. Si miramos la siguente grafica observamos que la tasa neta de incremento por individuo no se ve afectada por la competencia si N esta cerca de “0”. Cuando N aumenta hasta K, la tasa neta por individuo es cero. Teniendo en cuenta la linea A, B :
dN/dt(1/N)= (-r/K x N)=r que simplificando:
dN/dt(1/N)= r(1-N/K)
dN/dt= rN(K-N/K)
Estas ecuaciones han desarrollado un papel fundamental en el desarrollo de la ecologia. Es importante destacar tambien que se tratan de modelos muy limitados, pero he considerado sin profundizar en ello al menos representarlas.
7. Competencia interespecifica
7.1 definicion:
Al igual que ocurria con la competencia intraespecifica, se produce una disminucion de la tasa de natalidad, aumento de la mortalidad y una disminucion del indice de superivencia individual, debido a que los recursos que explota una determinada especie son tambien explotados por otras especies diferentes y a veces con mayor efectividad. Este hecho lleva a las especies a evitar explotar el mismo recurso para no solaparse y que la coexistencia sea posible. Sin embargo tambien se dan ejemplos en la naturaleza de exclusion de una especie por competencia si el recurso es lo suficientemente limitante.
7.2 Principio de la exclusion competitiva
Este principio puede resumirse en que dos especies no pueden coexistir indefinidamente sobre la base del mismo recurso limitante (ley del minimo de Liebig). Por lo que este principio predice que dos especies con iguales requerimientos no pueden existir. Sin embargo en la naturaleza se dan este tipo de coexistencias, pero como vamos a ver a continuacion esto es posible debido a que las diferencias aun siendo sutiles se dan.
7.3 Competencia en espacios desocupados
Cuando una especie excluye a otra es porque compiten en terminos de igualdad, la coexistencia es posible cuando cada especie se apropia de espacios diferentes (aunque tambien hemos visto que no es totalmente cierto). Esto es lo que puede ocurrir cuando en un ecosistema se produce una “perturbacion”. Tras una perturbacion (un incendio, un desprendimiento de tierra, etc.) el exito de los organismos que alli se instalen dependera en gran medida de lo vacantes que esten los nichos ecologicos que ocupen. Llegar primero tambien puede inclinar la balanza a la primera especie que lo consiga.
7.4 Competencia entre paridos:
El carbonero comun (Parus major), herrerillo (P. caeruleus), el carbonero palustre, el sibilino y el garrapinos (P. palustris, P. montanus y P. ater, respectivamente), son especies que pueden aparecer todas juntas en bosques de pinos y montaña. Todas estas aves nidifican en agujeros, todas comen insectos durante todo el año, todas buscan las mismas semillas durante el invierno y sin embargo, coexisten. Esto era posible porque siendo tan parecidas las especies de esta familia, habia algo que lo permitia, y es que cada ave estaba especializada en un estrato arboreo diferente, en un tamaño de insecto diferente y al parecer en una dureza de semilla diferente. Gracias a esto las especies coexisten porque explotan recursos ligeramente diferentes.
7.5 Competencia con diatomeas:
Este otro ejemplo ocurre justo lo contrario. Se trabajo con dos especies de diatomea de agua dulce: Asterionella formosa y Synedra ulna (Tilman et al.,1981). Como ya sabemos las diatomeas son algas unicelulares que requieren de silicatos para poder construir sus paredes celulares. Cultivando ambas especies por separado se observo que ambas poblaciones se mantuvieron estables. Sin embargo, Synedra consumia mas silicato que Asterionella, por lo que cuando ambas especies convivian Synedra dejaba los niveles de silicatos demasiado bajos como para que Asterionella pudiera sobrevivir.
7.6 Modelo matematico pera la competencia interespecifica:
Un modelo matematico que intente explicar como varian poblaciones con competencia interespecifica debe tener en cuenta que por lo que se compite es por los recursos, para ello existe un modelo logistico. El modelo de “Lotka y Volterra”
dN/dt= rN(K-N/K)
Ya hemos visto con anterioridad como este modelo explica los fenomenos de interacciones entre especies. En este caso el tamaño de la poblacion viene indicado por N1 y el de la segunda especie por N2. Sus capacidades de carga y sus tasas intrinsecas vienen indicadas por K1, y r1 respectivamente.
Para entender este modelo en la competencia interespecifica vamos a suponer que se dan dos especies A y B, de forma que al menos 10 individuos de la especie B ejerce el mismo grado de competencia que 1 individuo de la especie A. El efecto total que actua sobre la especie A sera equivalente al efecto de (N1+N2/10) individuos de la especie A. La constante 1/10 se llama coeficiente de competencia y se simboliza por α12.
De esta forma con estas dos expresiones se trabaja para el analisis de las especies A y B:
dN1/dt= r1N1(K1-N1- α12N2)/K1
Y en la segunda especie:
dN2/dt= r2N2(K2-N2- α21N2)/K1
Pero todo esto es mentira! las especies no evolucionan ni coevolucionan ni se adaptan ni interaccionas segun algunos...
Fuente BIBLIOGRAFIA Y REFERENCIAS
· Lovelock, Margulis, L. Bateson, G. Maturana, H. Varela, F. et all. (1989). “A way of knowing”. Edt. Kairós, S.A. Edt. The Lindisfarne Association
· M. Begon, J.L Harper, C.R.Towsend. (1999). “Ecology: Individuals, populations and comunities”. Tercera edición. Editorial Omega, S.A., Barcelona.
· Wilhelm Goestsch. (1987). “Die staaten der Ameisen”, La vida social de las hormigas. Segunda edición. Editorial Labor, S.A., Calabria.
· Edward E. Rupert & Robert D. Barnes. (1996). “Zoología de los Invertebrados”. Sexta edición. Editorial McGraw Hill Interamericana editores, S.A, México
· Universidad de Extremadura. (1994). “ETOLOGIA, introducción a las ciencias del comportamiento”. Tercera edición. Editorial Servicios de publicaciones de la universidad de Extremadura. Cáceres.
Apomios05 de jun. 2007Todo eso es paja, no respsonde a mis preguntas, que triste que tengas que recurrir a una biblia en verso pegada, que no aclara nada, sin poder dar tu opinion personal a mis preguntas
Es un trabajo de mi puño y letra...
Dejanos tranquilos.
Buen trabajo, Etpoursimouve.
Apomios05 de jun. 2007etpoursimouve
Hoy, a las 13:15
A nivel morfológico los cambios que se produjeron fueron inducidos por el hombre ya que la selección natural dejo de actuar sobre el lobo para actuar sobre el una nueva selección ( la artificial) en el lobo, la presión selectiva ejercida por el hombre origina nuevas tendencias de lo que iba buscando tanto a nivel morfológico como a nivel de comportamiento.
Apomios:
¿Entonces las razas de perros como dices no son evolucion y son una seleccion artificial?
¿Si encuentran un perro fosilizado en el futuro, diran que es evolucion o seleccion artificial?
Te dire algo, conozco muchos pastores que intentaron trabajar con cachorros de lobo para que hicieran la funcion de perros pastores y poder enfrentarse a los lobos salvajes con las mismas armas. Te dire lo que sucedio, hasta la fecha ninun cachorro de lobo educado con el hombre a podido cumplir la faena de un perro pastor, terminaban comiendose el ganado.
Apomios05 de jun. 2007Y digame señor bilogo
¿Cuantos miles de años segun usted, se necesitan para que un lobo iberico, se convierta por su seleccion artificial en un perro de aguas, un podenco o un alano español?
los mismos que necesitaría usted para entender algunos y elementales conceptos
Apomios05 de jun. 2007Su respuesta es la evidencia, me da la razón.
Apomios05 de jun. 2007Ya veo que cuando le formulan cuestiones, que escapan a sus esquemas mentales, la unica respuesta que tiene es salirse fuera del dialogo por los cerros hispanos.
Apomios05 de jun. 2007Tan difil es para usted contestar a mis preguntas sin faltarnos el respeto?
etpoursimouve
Hoy, a las 13:15
A nivel morfológico los cambios que se produjeron fueron inducidos por el hombre ya que la selección natural dejo de actuar sobre el lobo para actuar sobre el una nueva selección ( la artificial) en el lobo, la presión selectiva ejercida por el hombre origina nuevas tendencias de lo que iba buscando tanto a nivel morfológico como a nivel de comportamiento.
Apomios:
¿Entonces las razas de perros como dices no son evolucion y son una seleccion artificial?
¿Si encuentran un perro fosilizado en el futuro, diran que es evolucion o seleccion artificial?
¿Cuantos miles de años segun usted, se necesitan para que un lobo iberico, se convierta por su seleccion artificial en un perro de aguas, un podenco o un alano español?
Te dire algo, conozco muchos pastores que intentaron trabajar con cachorros de lobo para que hicieran la funcion de perros pastores y poder enfrentarse a los lobos salvajes con las mismas armas. Te dire lo que sucedio, hasta la fecha ninun cachorro de lobo educado con el hombre a podido cumplir la faena de un perro pastor, terminaban comiendose el ganado.
Le agradeceria, contestara de buenos modales mis preguntas
Gracias
¿Por qué los dioses (o el dios) crearon a los dinosaurios y luego los exterminaron? ¿Era algún tipo de experimento? ¿Se les castigo por no seguir las sagradas escrituras? ¿Aún no se le había ocurrido la idea de mamíferos bípedos? ¿se aburrían de verlos y decidieron crear una especie más interesante?¿O simplemente no existieron y sus huesos fósiles son todos falsos?
Espero alguien tenga la amabilidad de aclararme esa duda que tengo.
Un saludo
DeLaCuesta05 de jun. 2007Es una pregunta interesante la que formulaste Apomios, ¿Cuantos miles de años segun usted, se necesitan para que un lobo iberico, se convierta por su seleccion artificial en un perro de aguas, un podenco o un alano español?
A lo que añadiria, esta pregunta:
Cuantas mutaciones y con herencia genética posterior adquirida, tendrían que darse para ese cambio morfológico y de comportamiento, en cuantos miles de años se podría conseguir esa transmutación de un lobo a un perrillo falderillo, sin olvidarnos del gen de la herencia?
Si tienes la respuesta Apomios te agradecería que la escribieras y por supuesto a todos los demás, no me había planteado esas cuestiones, pero ahora que las he leido, me dejas con la incertidumbre, espero no me vuelvas loco.
No soy creacionista, pero me gustaría serlo, ahora que el 30% de los seres vivientes está al borde de la extinción, me reconfortaría.
Ello supera mi capacidad de comprensión; significaría que durante miles de millones de años, Potencias crearon y siguen creando, millones de millones de seres vivientes, desde priones, virus, algas, hasta mamíferos, insectos y etc. en todas sus variedades, inteligentemente adecuados a sus medios, hasta el momento de su desaparición, hechos todos con el mismo código genético, etc. etc.
Qué le vamos a hacer. Ansí es la vida.
UFFF esto del creacionismo es infumable, lo que no entiendo es como en el siglo XXI , haya gente inclinada a creer en estas cosas, y bueno lo de citar el corán (con todo mi respeto) para defenderlo ya es de traca de verdad.
Menos mal que gracias a muchos seguimos avanzando y conociendo más aún el planeta que pisamos, si fuera por otros , seguiríamos sin atravesar las columnas de hércules por miedo a encontrarnos con bestias marinas y un tremendo abismo sin fondo.
Algunos recordareis un "maravilloso" artículo que colgué en esta web, donde a raíz de el me topé con toda esta "gentecilla" que se dedica a intentar destrozar toda teoría científica actual (y sobradamente probada) a base de lanzar frases del corán y versitos iluminados....
En fín la religión y la mitología están muy bién , pero cuidado porque a algunos les ofusca la mente sobremanera..
Saúdos
Comparto el asombro y añado, que a mi me da un poco de miedo.
Apomios05 de jun. 2007Mas ofusca tu religion del mono.
y recuerda
“Nosotros os hemos creado. ¿Por qué pues no aceptáis la verdad? ¿Os habéis fijado en lo que eyaculáis? ¿Lo creáis vosotros o somos Nosotros los creadores?” (Corán 56:57-59).
Crees que estas palabras no son ciertas?
Apomios05 de jun. 2007Acepta la verdad y dejate de monsergas de monos voladores que se hacen hombres por arte de magia, cuanta imaginacion les diste a esta tribu Dios mio.
Jesús, Jesús, Jesús.
Lo terrible es que tu te las crees.
En, así nos va. Que depresión. Buaaaaaaaaaaaaaaaaa
Apomios05 de jun. 2007“Nosotros hemos construido el cielo con fuerza, y ciertamente lo estamos expandiendo” (Corán 51:47)
Como podia saber el profeta Mahoma (alabado sea su nombre), que el cielo se expande cuando se descubrio en el siglo XX?
Que si Apomios que si, aparte de mentar una y otra vez los mismos versos del corán , y cortapegas de otros foristas, me podías dar otra explicación a tu creacionismo? o saliéndote del corán no tienes más argumentos??
Religión del mono??? la religión es un acto de fé, tu lo deberías saber bien, la religión del mono que tu llamas, está basada en probadas teorías científicas, y si de algo tiene poco precisamente , es de religión, te lo vuelvo a decir, la religión si no la controlas acabará ofuscando tu mente, aunque en tu caso quizás no haya marcha atras ( y lo de la marcha atrás lo digo tb por tus eyaculaciones repetitivas )
"Los monos son demasiado buenos para que el hombre pueda descender de ellos."
"¿Es el hombre sólo un fallo de Dios, o Dios sólo un fallo del hombre?"
Friedrich Nietzsche
Y añado:
¿Por qué o para qué creó Dios a los dinosaurios?
Salud
Apomios05 de jun. 2007Solo dime una cosa
¿Como de la nada, se puede crear un sistema solar o el infinito, de donde salio la materia?
¿Como de la materia inerte sin vida, se puede crear vida inteligente?
Para que nos sintiéramos pequeñitos
Apomios05 de jun. 2007Y para que creo el ser humano?
En un tris!. ¿Diós no lo hizo en un solo día?
Hay 250 comentarios.
página anterior 1 2 3 4 ...5 página siguiente