Autor: ventero
viernes, 01 de junio de 2007
Sección: Artículos generales
Información publicada por: ventero
Mostrado 59.137 veces.
Los ancestros del hombre comenzaron a caminar primero en los árboles
Los primates que antecedieron al hombre moderno comenzaron a caminar en los árboles antes de hacerlo en suelo firme, según afirma un estudio de paleontólogos ingleses divulgado este jueves por la revista 'Science'.
De "El Mundo", Madrid, y EFE, Washington.-
Los primates que antecedieron al hombre moderno comenzaron a caminar en los árboles antes de hacerlo en suelo firme, según afirma un estudio de paleontólogos ingleses divulgado el 31 de mayo de 2007 por la revista 'Science'.

Las actuales teorías sobre la evolución de los homínidos señalaban que el bipedalismo, la característica que diferencia al hombre de sus parientes primates, comenzó cuando éstos descendieron de los árboles, no antes. Y una de las explicaciones más socorridas ha sido que los ancestros de chimpancés, gorilas y, por último, del hombre descendieron de los árboles para desplazarse en cuatro patas. Con el tiempo, comenzaron a caminar apoyándose en los nudillos y después únicamente sobre sus extremidades inferiores, como los seres humanos.
Sin embargo, según los paleontólogos de la Universidad de Liverpool, ese razonamiento comenzó a complicarse cuando estudios recientes hechos a partir de fósiles mostraron que algunos homínidos, incluido el de Lucy (australopithecus afarensis), vivieron en bosques.

Al mismo tiempo, otras formas más primitivas como la del Hombre del Milenio (Orrorin) parece que vivieron en la copa de los árboles y se desplazaban sobre sus extremidades inferiores. "Los resultados de nuestro estudio han confundido el panorama", señaló Robin Crompton, uno de los autores de la investigación. "Si estamos en lo cierto, esto significa que no se puede confiar en que el bipedalismo nos diga si estamos ante el ancestro de un ser humano o el de otro primate", indicó. Según el científico, "es cada vez más difícil decir qué es humano y qué es simio y nuestro trabajo lo confirma".
Crompton, Susannah Thorpe y Roger Holder, de la Universidad de Birmingham, llegaron a la conclusión sobre el bipedalismo arbóreo de los primates al observar orangutanes en Sumatra (Indonesia). Según señalaron, los orangutanes pasan la mayor parte de su vida en los árboles, lo que los convierte en modelos útiles para establecer cómo se desplazaron nuestros ancestros hace millones de años.
Debido a que esos ancestros comían frutas, habrían tenido que apoyarse en las ramas débiles en la periferia de la copa de los árboles. El hecho de caminar sobre sus extremidades inferiores y utilizar sus brazos para mantener el equilibrio les habría ayudado a desplazarse, según el estudio. "Nuestros resultados sugieren que el bipedalismo se usó para recorrer las ramas más débiles, donde están las mejores frutas, y también para pasar de un árbol a otro", según Thorpe.
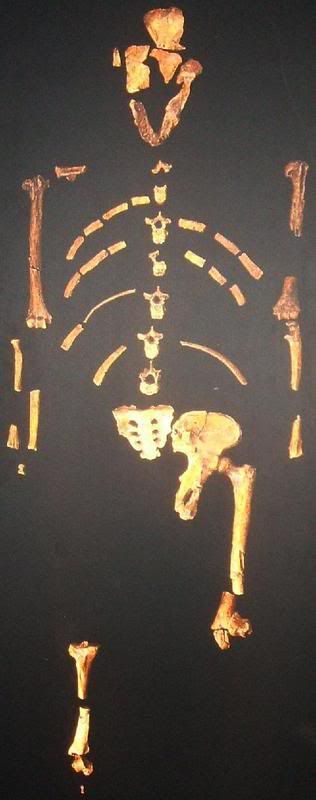
Los científicos creen que al bipedalismo contribuyó un cambio climático registrado entre 5 y 24 millones de años atrás, cuando disminuyó la frondosidad de los árboles. En un ambiente más árido, comenzaron a encontrar dificultades para pasar de un árbol a otro debido a la disminución de la espesura. Los ancestros humanos respondieron a este problema abandonando las copas de los árboles para llegar al suelo de las selvas, donde mantuvieron su bipedalismo y comenzaron a alimentarse en árboles más pequeños. Por su parte, los ancestros de chimpancés y gorilas se especializaron más en el ascenso vertical de los árboles y desarrollaron su forma de caminar sobre los nudillos.
"Nuestra conclusión es que el bipedalismo arbóreo tuvo grandes beneficios adaptativos. Por ello no necesitamos explicar cómo nuestros ancestros pasaron de ser cuadrúpedos a bípedos", dijo Thorpe.
-
No hay imágenes relacionadas.
Comentarios cerrados por el administrador Silberius
Motivo: Degradación en la calidad de los comentarios
Comentarios
 Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Tu único clic no la borarrá, pero contribuirá a que la sabiduría del grupo pueda funcionar correctamente.
Comentarios cerrados por el administrador Silberius
Motivo: Degradación en la calidad de los comentarios
Apomios04 de jun. 2007“Os ha creado en los vientres de vuestras madres, etapa tras etapa (creación tras creación), en una tRIPle oscuridad. Tal es Dios, vuestro Señor. Suya es la Soberanía. No hay divino sino El. ¡Cómo podéis, pues, ser tan desviados!” (Corán 39:6).
Apomios05 de jun. 2007Teoría científica
Planteamiento de un sistema abstracto hipotético-deductivo que constituye una explicación o descRIPción científica a un conjunto relacionado de observaciones o experimentos. Así, una teoría científica está basada en hipótesis o supuestos verificados por grupos de científicos (en ocasiones un supuesto, no resulta directamente verificable pero sí la mayoría de sus consecuencias). Abarca en general varias leyes científicas verificadas y en ocasiones deducibles de la propia teoría. Estas leyes pasan a formar parte de los supuestos e hipótesis básicas de la teoría que englobará los conocimientos aceptados por la comunidad científica del campo de investigación y está aceptada por la mayoría de especialistas.
http://es.wikipedia.org/wiki/Teor%C3%ADa
Como ven, teoria es algo que no se puede demostrar, solo hipotesis y si no estan deacuerdo hablen con el wikipedia.
Apomios05 de jun. 2007HARUN YAHYA
http://www.harunyahya.com/es/milagros_02a.php
Extraido de milagros del Coran
EL COLAPSO CIENTÍFICO DEL DARWINISMO
Aunque se trata de una doctrina que se remonta en el pasado hasta la Grecia Antigua, la teoría de la evolución avanzó considerablemente sobre todo en el siglo XIX. El trabajo más importante, que hizo que la teoría se convirtiera en el tema principal del mundo científico, fue el libro de Charles Darwin titulado “El origen de las especies” publicado en 1859. En este libro Darwin niega que las diferentes especies vivas que habitan o habitaron la tierra fueran creadas separadamente por Dios. Según Darwin, todos los seres vivientes provienen de un ancestro común y se han diversificado a lo largo del tiempo a través de pequeños cambios.
La teoría de Darwin no está basada en ningún descubrimiento científico concreto; como incluso él lo aceptó, no es más que una “presunción”. Más aún, como Darwin confiesa en el largo capítulo de su libro titulado “Dificultades de la teoría”, su hipótesis fallaba al momento de tratar de explicar muchas cuestiones críticas.
Darwin ponía toda su esperanza en los nuevos descubrimientos científicos, que esperaba resolvieran esas “dificultades de la teoría”. Sin embargo, contrariando sus expectativas, los descubrimientos científicos posteriores expandieron las dimensiones de esas dificultades.
La derrota del darwinismo por la ciencia se puede sintetizar en tres tópicos básicos:
1) La teoría no puede explicar por ningún medio cómo se originó la vida sobre la tierra.
2) No existen hallazgos científicos que muestren que los “mecanismos evolutivos” propuestos por la teoría tengan algún poder para provocar la evolución.
3) Los restos fósiles prueban exactamente lo contrario de lo que sugiere la teoría de la evolución.
LA PRIMERA ETAPA INSUPERABLE: EL ORIGEN DE LA VIDA
La teoría de la evolución postula que todas las especies vivas evolucionaron de una única célula que surgió en la tierra primitiva hace 3.800 millones de años. Cómo una única célula pudo generar millones de complejas especies vivas y, si tal evolución realmente ocurrió, porqué no pueden observarse vestigios de ella en los restos fósiles, son algunas de las cuestiones que la teoría no puede responder. Empero, y por encima de todo, en la primera etapa del alegado proceso evolutivo, habría que preguntarse: ¿cómo se originó esa “primera célula”?
Dado que la teoría de la evolución niega la creación y no acepta ningún tipo de intervención sobrenatural, sostiene que la “primera célula” fue un resultado casual de las leyes naturales sin ningún proyecto, plan u orden. Según la teoría la materia inanimada debió haber producido una célula viva por casualidad. Pero sin embargo esta es una pretensión inconsistente, incluso, con las más inconmovibles reglas de la biología.
“LA VIDA PROVIENE DE LA VIDA”
En su libro Darwin jamás se refiere al origen de la vida. El primitivo conocimiento científico que se tenía en su época descansaba en la suposición de que los seres vivientes tenían una estructura muy simple. Desde épocas medievales estaba ampliamente difundida la generación espontánea, una teoría que afirma que la materia inanimada puede producir directamente seres vivos. Estaba ampliamente difundida la creencia de que los insectos eran producidos por las sobras de comida, y los ratones por el trigo. Se realizaban interesantes experimentos para probar esta teoría. Se colocaba algo de trigo sobre una pieza sucia de tela y se creía que de ello se originarían ratones pasado un tiempo. De manera análoga los gusanos que aparecían en la carne se consideraban como una evidencia de la generación espontánea. Pero tiempo después se comprendió que los gusanos no aparecían sobre la carne espontáneamente, sino que surgían de las larvas depositadas allí por las moscas y que eran invisibles a ojo desnudo.
Incluso en la época en que Darwin escribió “El origen de las especies” la creencia de que las bacterias podían generarse de materia inerte era todavía ampliamente aceptada en el mundo científico. Pero cinco años después de que el libro de Darwin fuera publicado el descubrimiento de Luis Pasteur echó por tierra esta creencia, que constituía el fundamento de la evolución. Pasteur resumió la conclusión a la que había arribado después de largos estudios y experimentos con esta frase: “La hipótesis de que la materia inanimada puede originar la vida ha quedado enterrada en la historia para siempre” (44).
Los defensores de la teoría de la evolución resistieron los hallazgos de Pasteur durante largo tiempo. Pero a medida que la ciencia avanzaba y desentrañaba la compleja estructura de la célula de un ser viviente, la idea de que la vida podía producirse por azar se enfrentó con un atolladero aún mayor.
ESFUERZOS QUE NO CONVENCEN EN EL SIGLO XX
El primer evolucionista que retomó el tema del origen de la vida en el siglo XX fue el famoso bioquímico ruso Alexander Oparin. Con varias tesis en las que trabajó durante la década de 1930 trató de probar que la célula de un ser viviente podía originarse por azar. Estos estudios, sin embargo, estaban condenados al fracaso, y Oparin tuvo que hacer la siguiente confesión: “Desafortunadamente, el origen de la célula continúa siendo el punto más oscuro de toda la teoría de la evolución” (45).
Algunos evolucionistas seguidores de Oparin trataron de llevar a cabo experimentos para resolver el problema del origen de la vida. El más conocido de estos experimentos fue el que realizó el químico americano Stanley Miller en 1953. Combinando gases que sostenía existían en la atmósfera primordial de la tierra en un balón de vidrio, y bombardeándolos con energía eléctrica, Miller sintetizó varias moléculas orgánicas (aminoácidos) presentes en la estructura de las proteínas.
Habían pasado escasamente unos pocos años de este experimento —que era presentado como un paso importante para demostrar la teoría evolucionista— cuando se reveló que el mismo era inválido pues la atmósfera utilizada en el mismo era muy diferente de las condiciones reales existentes en la tierra (46).
Después de un largo silencio, Miller confesó que el medio atmosférico que había utilizado era irreal (47).
Todos los esfuerzos evolucionistas emprendidos durante el siglo XX para explicar el origen de la vida terminaron en fracaso. El geoquímico Jeffrey Bada del San Diego ScRIPps Institute, acepta esto en un artículo publicado en la revista Earth en 1998:
“Hoy, terminando el siglo XX, enfrentamos todavía el mayor problema sin resolver que teníamos cuando comenzó el siglo: ¿cómo se originó la vida sobre la tierra?” (48).
LA COMPLEJA ESTRUCTURA DE LA VIDA
La razón fundamental por la cual la teoría de la evolución terminó en semejante atolladero en lo referente al origen de la vida se debe a que incluso los organismos vivientes que se supone más simples tienen estructuras increíblemente complejas. La célula de un ser viviente es más compleja que todos los productos tecnológicos producidos por el hombre. Actualmente, incluso en los laboratorios más modernos del mundo, es imposible producir una célula viva uniendo materia inorgánica.
Las condiciones requeridas para la formación de una célula son cuantitativamente demasiado grandes para ser explicadas por la casualidad. La probabilidad de que las proteínas, principales componente de las células, resulten sintetizadas por casualidad es de 1 en 10.950 para una proteína promedio compuesta de unos 500 aminoácidos. En matemáticas una probabilidad menor a 1 en 1050 es considerada prácticamente un imposible.
La molécula de ADN, que está ubicada en el núcleo de la célula y que almacena la información genética, es una base de datos increíble. Se calcula que si la información codificada en el ADN se pusiera por escrito, ello daría lugar a una inmensa biblioteca equivalente a una enciclopedia de 900 volúmenes de 500 páginas cada uno.
Y en este punto aparece un dilema muy interesante: el ADN sólo puede replicarse con la ayuda de algunas proteínas especiales (enzimas). Pero la síntesis de estas enzimas sólo puede realizarse a partir de la información codificada en el ADN. Como ambas dependen una de otra tienen que existir al mismo tiempo para replicarse. Esto lleva a un punto muerto al escenario en el cual la vida se origina por sí misma. El profesor Leslie Orgel, un reputado evolucionista de la Universidad de San Diego, California, confesó este hecho en la edición de septiembre de 1994 de la revista “Scientific American”:
“Es extremadamente improbable que las proteínas y los ácidos nucleicos, ambos estructuralmente complejos, surjan espontáneamente en el mismo lugar al mismo tiempo. Además, parece también imposible obtener uno sin el otro. Y en consecuencia, a primera vista, uno tendría que concluir que la vida, en realidad, nunca pudo originarse por medios químicos” (49).
Indudablemente, es imposible que la vida se haya originado por causas naturales, y entonces no queda sino aceptar que fue “creada” por medios sobrenaturales. Este hecho invalida explícitamente la teoría de la evolución cuyo propósito principal es negar la creación.
LOS MECANISMOS IMAGINARIOS DE LA EVOLUCIÓN
El segundo tópico importante que invalida la teoría darwinista es que los dos conceptos expuestos por ésta como “mecanismos evolutivos” se ha visto que, en realidad, no poseen ningún poder evolutivo.
Darwin basó su alegato en favor de la evolución enteramente en el mecanismo de la “selección natural”. La importancia que le adjudica a este mecanismo queda en evidencia por el título mismo de su libro: “El origen de las especies por medio de la selección natural”.
La selección natural sostiene que aquellos seres vivientes que son más fuertes y están más adaptados a las condiciones naturales de su hábitat sobrevivirán en la lucha por la vida. Por ejemplo, en una manada de ciervos amenazada por animales salvajes, aquellos que puedan correr más rápido sobrevivirán. En consecuencia, la manada de ciervos estará compuesta de los individuos más fuertes y más rápidos. Sin embargo, incuestionablemente, este mecanismo no causará que los ciervos evolucionen y se transformen en otra especie, por ejemplo, caballos.
Consecuentemente el mecanismo de la selección natural no tiene poder evolutivo. Darwin era consciente de este hecho y lo dejó expresado en su libro “El origen de las especies”:
“La selección natural no puede hacer nada hasta que ocurran variaciones favorables fortuitas” (50).
EL IMPACTO DE LAMARCK
Ahora bien, ¿cómo pueden ocurrir estas “variaciones favorables”? Darwin trató de responder a esta cuestión partiendo del punto de vista que el primitivo estado del conocimiento científico poseía en su época. Según el biólogo francés Lamarck, que vivió antes de Darwin, las criaturas vivas transmitían los rasgos que adquirían durante su vida a la siguiente generación, y estos rasgos acumulándose de una generación a otra provocaban la aparición de nuevas especies. Por ejemplo, según Lamarck, las jirafas evolucionaron de los antílopes; esforzándose por comer hojas de árboles altos sus cuellos fueron estirándose de una generación a otra.
Darwin da también ejemplos similares en su libro “El origen de las especies”. Por ejemplo, dice que algunos osos que se introducían en el agua para buscar comida se transformaron en ballenas con el paso del tiempo (51).
Pero no obstante, las leyes de la herencia descubiertas por Mendel y verificadas por la genética que floreció en el siglo XX, finalmente destruyeron la leyenda de que los rasgos adquiridos se trasmitían a las generaciones subsiguientes. De esta forma la selección natural perdió sustento como mecanismo evolutivo.
EL NEODARWINISMO Y LAS MUTACIONES
Para encontrar una solución, los darwinistas propusieron a finales de la década de 1930 la “moderna teoría sintética” o, como se la conoce más comúnmente, el neodarwinismo.
El neodarwinismo agregó las mutaciones, que son distorsiones producidas en los genes de los seres vivientes como resultado de factores externos tales como radiaciones o fallas en la replicación, como “causa de las variaciones favorables” además de la selección natural.
Actualmente el modelo evolucionista que persiste en el mundo es el neodarwinista. La teoría sostiene que los millones de seres vivientes presentes en la tierra son el resultado de mutaciones o desórdenes genéticos por medio de los cuales se fueron provocando cambios en numerosos órganos complejos de estos organismos, tales como oídos, ojos, extremidades, alas, etc. Sin embargo, existe un hecho científico innegable que socava esta teoría: las mutaciones no provocan una evolución en los seres vivientes; por el contrario siempre los perjudican.
La razón para esto es muy simple: el ADN tiene una estructura muy compleja y las mutaciones azarosas sólo pueden dañarla. El genetista americano B. G. Ranganathan explica esto como sigue:
“Las mutaciones son pequeñas, azarosas y dañinas. Ocurren raramente y lo mejor que puede pasar es que sea ineficaz. Estas cuatro características de las mutaciones implican que no pueden conducir a un desarrollo evolutivo. Un cambio al azar en un organismo altamente especializado o es dañino o es fútil. Un cambio al azar en un reloj pulsera no puede mejorarlo. Lo más probable es que lo perjudique o, en el mejor de los casos, que sea insustancial. Un terremoto no mejora una ciudad, la destruye” (52).
No es sorprendente que ningún ejemplo útil de mutación, esto es, que se haya constatado que mejoró el código genético, se haya observado hasta ahora. Todas las mutaciones han probado ser nocivas. La mutación, que se presenta como un “mecanismo evolutivo”, es realmente un incidente genético que daña a los seres vivientes y los incapacita. (El efecto más común de mutación en los seres humanos es el cáncer). No caben dudas que un mecanismo destructivo no puede ser un “mecanismo evolutivo”. La selección natural, por otro lado, “no puede hacer nada por sí misma” como también Darwin aceptó. Esto nos indica que no hay “mecanismos evolutivos” en la naturaleza, y si no existen difícilmente alguien pueda imaginar cómo el proceso llamado evolución tendría lugar.
RESTOS FÓSILES: NO HAY RASTROS DE FORMAS INTERMEDIAS
La prueba más clara de que el escenario sugerido por la teoría evolucionista no existe son los restos fósiles.
Según la teoría de la evolución todas las especies han surgido de una precedente. Es decir que especies previas sufrieron transformaciones a lo largo del tiempo, y todas se produjeron de este modo en un proceso gradual de transformación que duró millones de años.
Si éste ha sido el caso, entonces debieron existir numerosas especies intermedias que vivieron durante este larguísimo período de transformación.
Por ejemplo, alguna especie medio-pez/medio-reptil debió haber vivido en el pasado adquiriendo, con el paso del tiempo, algunas características de reptil además de las de pez que ya tenía. O debieron existir algunos reptiles-pájaros que adquirieron más características de las aves aparte de las de reptil que ya poseían. Dado que estas especies estaban en una fase de transición, debía tratarse de seres vivos defectuosos, limitados por ciertas incapacidades. Los evolucionistas se refieren a estos seres imaginarios, que ellos creen que vivieron en el pasado, como “formas transitorias”.
Si tales animales realmente han existido, deberían haber sido millones, o incluso miles de millones en cuanto a su número y variedad. Y más importante todavía, los restos de estos extraños seres deberían estar presentes en los restos fósiles. En “El origen de las especies” Darwin explicaba:
“Si mi teoría es cierta, innumerables variedades intermedias, como eslabones cercanos de todas las especies de un mismo grupo, ciertamente deben haber existido... Consecuentemente, evidencia de su existencia previa sólo podría hallarse entre los restos fósiles” (53).
LAS ESPERANZAS DE DARWIN SE HICIERON PEDAZOS
Pese a que los evolucionistas han realizado enérgicos esfuerzos en todo el mundo para encontrar fósiles desde mediados del siglo XIX, todavía no se han descubierto formas intermedias. Todos los fósiles desenterrados en las excavaciones muestran que, contrariamente a las expectativas de los evolucionistas, todas las formas de vida aparecieron sobre la tierra en forma repentina y completamente formadas.
Un paleontólogo británico, Derek V. Ager, admite este hecho aunque él es un evolucionista:
“La cuestión que surge es que, si nosotros examinamos en detalle los restos fósiles, sea a nivel de órdenes o de especies, encontramos —una y otra vez— no una evolución gradual, sino una explosión repentina de un grupo a expensas de otro” (54).
Esto significa que en los restos fósiles todas las especies surgen repentinamente y completamente formadas, sin ninguna forma intermedia en el medio. Esto es exactamente lo opuesto de las presunciones de Darwin. Y además es una evidencia muy fuerte de que los seres vivientes son creados. La única explicación para que una especie viviente aparezca repentinamente y completa en todos sus detalles, sin ningún ancestro del cual haya evolucionado, es que fue creada. Este hecho es admitido también por el ampliamente conocido biólogo evolucionista Douglas Futuyma:
“La creación y la evolución, entre ambas, agotan todas las explicaciones posibles para el origen de los seres vivientes. Los organismos vivos o bien aparecen sobre la tierra completamente desarrollados o no lo hacen. Si no lo hacen deben haber evolucionado de especies preexistentes por algún proceso de modificación. Y si aparecen en un estado completamente desarrollado, deben haber sido creados por alguna inteligencia omnipotente” (55).
Los fósiles muestran que los seres vivientes aparecieron sobre la tierra completamente desarrollados y en un estado perfecto. Esto significa que “el origen de las especies”, contrariamente a lo que suponía Darwin, no es la evolución sino la creación.
EL CUENTO DE LA EVOLUCIÓN HUMANA
El tema que traen a colación más a menudo los defensores de la teoría evolucionista es el del origen del hombre. La tesis darwinista sostiene que el hombre moderno actual evolucionó de algún tipo de criatura simiesca. Durante este supuesto proceso evolutivo, que se supone comenzó hace 4-5 millones de años, se afirma que han existido algunas “formas de transición” entre el hombre moderno y sus ancestros. Según este escenario completamente imaginario, existen cuatro “categorías” básicas:
1. Australopithecus
2. Homo habilis
3. Homo erectus
4. Homo sapiens
Los evolucionistas llaman “Australopithecus” al primero de estos ancestros similares a los simios, palabra que significa “simio sudafricano”. Esos seres no eran en realidad más que una antigua especie de simios que se ha extinguido. Profundos estudios realizados sobre varios especímenes del Australopithecus por dos anatomistas mundialmente famosos de Inglaterra y EE.UU., Lord Solly Zuckerman y el Prof. Charles Oxnard, han mostrado que esos fósiles pertenecen a una especie ordinaria de simio que se ha extinguido y que no presenta semejanzas con los seres humanos (56).
Los evolucionistas clasifican a la siguiente etapa de la evolución humana como “homo”, es decir “hombre”. Según sus afirmaciones las criaturas de la serie “homo” están más desarrolladas que el Australopithecus. Pero lo que hacen es inventar un esquema evolutivo imaginario ordenando diferentes fósiles de esas criaturas en un cierto orden. Este esquema es imaginario porque jamás se ha probado que exista una relación evolutiva entre estas diferentes clases. Ernst Mayr, uno de los principales defensores de la teoría de la evolución en el siglo XX, admite este hecho diciendo que “la cadena que llega hasta el homo sapiens está en realidad perdida” (57).
Delineando la cadena de eslabones en la forma “Australopithecus > Homo habilis > Homo erectus > Homo sapiens” los evolucionistas dan a entender que cada una de estas especies es ancestro de la siguiente. Pero sin embargo, recientes descubrimientos de los paleoantropólogos han revelado que el Australopithecus, el Homo habilis y el Homo erectus han vivido en diferentes partes del mundo al mismo tiempo (58).
Más aún, ciertos segmentos de humanos clasificados como Homo erectus han vivido hasta épocas muy modernas. El Homo sapiens neanderthalensis y el Homo sapiens sapiens (el hombre moderno) coexistieron en la misma región (59).
Esta situación claramente indica la invalidez de la hipótesis que sostiene que son ancestros unos de otros. Un paleontólogo de la Universidad de Harvard, Stephen Jay Gould, explica este punto muerto de la teoría de la evolución, aunque él mismo es un evolucionista, en estos términos:
“¿Qué ha pasado con nuestra escalera si existen tres linajes de homínidos coexistentes (australopithecus africanus, el robusto australopithecus, y el homo habilis), ninguno claramente derivado del otro? Más aún, ninguno de los tres muestra tendencias evolutivas durante su estancia en la tierra” (60).
En resumen, el escenario de la evolución humana que se presenta en los medios de comunicación y en los textos escolares apoyado en varios dibujos de algunas criaturas “mitad simios, mitad humanos”, es, francamente hablando, simple propaganda, pues no es otra cosa que un cuento sin ningún fundamento científico.
Lord Solly Zuckerman, uno de los más famosos y respetados científicos del Reino Unido, que llevó a cabo investigaciones sobre este tema durante mucho tiempo, y que en particular estudió los fósiles del Australopithecus durante 15 años, llegó finalmente a la conclusión —pese a que él es un evolucionista— de que, en realidad, no existe ninguna ramificación evolutiva que, partiendo de esas criaturas parecidas a los simios, termine en el hombre.
Zuckerman es autor además de una interesante “clasificación de la ciencia”. Elaboró un cuadro jerárquico de las disciplinas científicas ordenándolas desde las que él considera científicas hasta las que considera a-científicas. Según la clasificación de Zuckerman, los campos de la ciencia más “científicos” —es decir, dependientes de datos concretos— son la química y la física. Después de ellos vienen las ciencias biológicas y luego las ciencias sociales. Al final de la tabla, que es la parte considerada más “a-científica”, están la “percepción extrasensorial” —temas tales como la telepatía y el sexto sentido— y finalmente la “evolución humana”. Zuckerman explica así su razonamiento:
“Nos desplazamos entonces fuera del registro de las verdades objetivas para entrar en el campo de la ciencia biológica presuntiva, como la percepción extrasensorial o la interpretación de la historia fósil del hombre, donde para el convencido (evolucionista) todo es posible, y donde el ardiente creyente (en la evolución) es algunas veces capaz de creer varias cosas contradictorias al mismo tiempo” (61).
Copio y pego un extracto de un trabajo de Ecologia
Dinamica y efectos de la predación
2.1 Efectos:
Estos efectos suelen ser perjudiciales en la poblacion de presas, pero existen casos que si son perjudiciales para la presa no lo son tanto para el conjunto de la poblacion, esto se debe a que si la densidad de la poblacion de presas es lo suficientemente alta como para que se esten dando fenomenos de competencia intraespecifica, el efecto de la depredacion hara efectiva una reduccion del numero de individuos que compiten por el alimento o el espacio. Por esto la competencia intraespecifica se vera aliviada.
De cualquier forma los depredadores van a determinar en ultima instancia la abundancia de las presas, o viceversa. Para ello existe un modelo matematico que intenta explicar como funcionan este tipo de interacciones llamado Lotka – Volterra
2.2 Modelo de Lotka y Volterra:
Recibe su nombre de los que fueron sus creadores, es un modelo que posee dos componentes; C, el numero de individuos existentes en una poblacion de consumidores, y N, que es el numero de individuos existentes en la poblacion de presas.
De este modo se puede decir que en ausencia de consumidores las presas aumentaran su poblacion exponencialmente:
dN∕dt=rN
sin embargo los individuos presa seran eliminados con una frecuencia que dependera de los encuentros que se den entre la presa y su consumidor, esta frecuencia aumentara en funcion del aumento de depredadores C, yo presas N.
Los encuentros fuctiferos y la tasa de consumo seran:
dN∕dt=rN-aCN
Donde aCN es la tasa de consumo. Este modelo tambien explica que en ausencia de alimento los depredadores disminuiran su eficacia y moriran de hambre, por lo que su numero decrecera exponencialmente:
dC∕dt=-qC
Donde q es la tasa de mortalidad. Pero esto se ve contrarrestado por el nacimiento de nuevos depredadores, cuya tasa depende de:
i) La tasa de consumo de alimento
ii) Eficacia, f, para convertir el alimento en descendientes
Por lo que la tasa de natalidad es faCN
dC∕dt=faCN-qC
Este modelo resulta muy util para señalar las tendencias en las relaciones depredador presa, de forma que se estudian las oscilaciones entre ambos linajes. Este mecanismo de oscilaciones acopladas posee una serie de retrasos en el tiempo, esto se debe a que los periodos de abundancia de presas no dan tiempo a satisfacer a la poblacion de presas. De este modo se produce un retraso en el tiempo de pocas presas y pocos depredadores y asi sucesivamente.
3. Preferencias alimentincias
3.1 Preferencias y tipos de consumidores:
Existen diferentes modelos de comportamiento en los consumidores ala hora de elegir o seleccionar aquello que va a ser consumido.
En lineas generales vamos a encontrar 3 tipos de consumidores. Monofagos (solo consumen un tipo de especie), olifagos ( consumen solo varias especies), polifago (consumen un rango muy alto de especies). Es decir, las especies monofagas seria las tambien llamadas especialistas, las polifagas son las generalistas y las olifagas serian las que tienen un rango intermedio.
Sin embargo las especies denominadas “generalistas” no toman cualquier cosa que se encuentren como alimento. Sino que aceptan solo una estrecha gama de los recursos existentes ( un raton es un recurso pero un leon no se lo come). Es decir, dentro de la generalidad de una especie polifaga u olifaga, existe una preferencia a la hora de elegir alimento.
Estas preferencias estan siempre sujetas a lo provechosas que puedan llegar a resultar para el individuo que las consumen, dicho de otro modo: Los individuos consumidores tenderan a escoger las presas mas energeticas.
Pero esto no siempre resulta asi, en la alimentacion de los consumidores herbivoros muchas veces la eleccion del alimento esta condicionada la concentracion de las toxinas que las plantas pueda contener y esto puede llevar en ambientes donde las plantas estan bien equipadas a la herbivoria a una dieta mixta. En estos casos una dieta mixta mantiene las concentraciones de toxinas a niveles adecuados, ya que cada especie vegetal sintetiza una toxina propia.
3.2 Amplitud de la dieta:
El primer trabajo que desarrolla un modelo en la amplitud de la dieta fue el llamado teoria del aprovisionamiento optimo (Mc Arthur & Pianka). En esta teoria se argumenta que para obtener energia todo depredador necesita invertir tiempo y energia, primero en buscarla y luego en manipularla, por ello los investigadores consideraron que la amplitud de la dieta dependia en ultima instancia de la capacidad de manipulacion, de esta forma cuando un depredador generalista busca presas encuentra un alto numero, mientras que un especialista solo consume la presa que prefiere. Esto plantea un gran peoblema si el individuo es un especialista, dado que solo consumira presas provechosas, los tiempos de busqueda pueden resultar altos y puede invertir mucho tiempo y energia. Mientras, si es un generalista los tiempos de busqueda seran breves.
Segun esto la teoria hace 4 predicciones:
i) Los buscadores deberian ser generalistas
ii) Los manipuladores especialistas
iii) La especializacion deberia ser mayor en ambientes productivos
iv) La abundancia de los tipos de presas no provechosas es irrelevante
La primera de estas afirmaciones parece verse confirmada si tenemos en cuenta a las especies de aves paseriformes insectivoras como la curruca cabecinegra Sylvia melanocefala, cuya dieta esta compuesta esencialmente por diminutas larvas de insetos, esto requiere un tiempo muy corto de manipulacion. Por lo que una dieta amplia le conlleva mas ventajas que desventajas. Por el contrario depredadores como el leon africano Phantera leo, viven casi la totalidad de su vida a la vista de sus presas,
por lo que el gasto en encontrarlas es casi nulo e invierten mas en la manipulacion, la persecucion puede llegar a ser mas larga que la eleccion de la presa y por tanto se especializan en las presas provechosas (enfermas, jovenes y viejos).
3.3 Carrera de armamentos:
De cualquier forma la polifagia parece ofrecer mayores ventajas. La inversion en busqueda es menor, morir de hambre es mas dificil y aun asi se dan en la naturaleza casos en los que asociaciones depredador-presa lejos de ampliar las dietas se han visto reducidas a la minima expresion, es decir una planta o presa es consumida por un solo tipo de herbivoro o depredador, y a su ven estos solo pueden consumirlos a ellos. Esto solo se entiende en el contexto de la evolucion y en este caso las parejas depredador presa no solo han evolucionado si no que han coevolucionado. La idea es que se produce una “carrera de armamentos” evolutiva en la que existe una mejora progresiva en la efectividad de caza de un predador para con una presa cualquiera pero a su vez esta tambien va mejorando su capacidad de escape. Esta idea nos sugiere que si trasladasemos un predador de ratones del Eoceno quizas se moriria de hambre intentando capturar a uno actual, y ademas nos explica porque ciertos insectos se parecen a hojas o la semejanza entre una polilla y una corteza de arbol, y esto es probable gracias a una larga carrera de armamentos entre insectos y aves insectivoras dispuestas a discriminar entre las verdaderas presas y sus modelos no comestibles.
3.3.1 Factores que otorgan ventajas:
A veces se dan situaciones donde algunas de la partes implicadas juega con ventajas. Sin que con ello se quiera decir que esta parte de la contienda vaya a ganar la carrera de armamentos, lo que ocurre es que una de las partes experimenta procesos de evolucion mas altos que la otra. Cada vez que uno de los dos linajes experimente una adaptacion, disfrutara de un periodo de relajacion ya que el otro linaje disminuira su eficacia durante algun tiempo. Existen varios factores que otorgan ventajas a una de las partes:
i) Las presiones selectivas son desiguales. Para una presa y un depredador las presiones selectivas son desiguales ya que uno corre por su vida mientras que el otro lo hace por su cena. Esto significa que el conejo que no escape a su perseguidor jamas se reproducira, mientras que el zorro si lo hara aunque equel dia no llegase a capturar el conejo. Por ello la asignacion del conejo a “escapar del zorro” sera mayor que la del zorro “atrapar conejo”, por lo que la eficacia global es mayor en las presas que en los depredadores y parten siempre con una ligera ventaja.
ii) Efecto enemigo raro. Aqui uno de los dos linajes explota al otro limpiamente, y lo hace con el efecto del engaño. En las luciernagas, los machos y las hembras se reunen para la reproduccion. Algunas luciernagas del genero Photuris imitan las señales de las hembras del genero Photinus de manera que cuando los machos se acercan para un posible cortejo, estos son capturados y devorados. Pero podriamos preguntarnos por que estas se dejan engañar por las “femme fatale”, la razon es simple, el depredador es una especie mas rara que la presa.
iii) Presupuestos adaptativos. Cualquier adaptacion es costosa. Si un linaje se centra en una carrera de armamentos concreta obviara otros asuntos, un Guepardo puede evolucionar hacia una carrera aun mas rapida pero obviara quizas la maniobrabilidad y la resistancia para arrastrarla y consumirla.
iv) Tasas adaptativas. Segun el registro fosil, los diferentes grupos taxonomicos no han evolucionado a la misma velocidad. Una salamandra actual se parece mas a un anfibio de Carbonifero que un ave actual a un Archaeopteryx, y ya sabemos que estas dos ultimas estan mas cerca en el tiempo. Los factores que pueden acelerar estos procesos son muy diversos; varianza genetica , aislamiento poblacional, tiempo de generacion cortos, etc, etc.
4. Parasitismo, parasitos y parasitoides
4.1 Clasificacion y definicion:
Como ya se definio en el apartado 1.3, los parasitos son organismos que consumen solo una parte de los recursos de un organimo, suelen causarles daño y a veces la muerte. Sin embargo su clasificacion no ha sido tan sencilla como su definicion, asi que tomaremos la diferenciacion entre micro y macroparasitos, de forma general:
i) Los microparasitos son pequeños, muy numerosos, y se multiplican en el interior de sus huespedes. La mayoria de estos son intracelulares y son los responsables de las infecciones bacterianas y viricas. GRIPe, malaria, y sarampion son algunos ejemplos.
ii) Los macroparasitos son bastante diferentes a los anteriores, generalmente poseen una fase de crecimiento en el interior del huesped, y otra de multiplicacion en el exterior, donde la descendencia busca nuevos hospedadores que parasitar. Piojos, pulgas, garrapatas, duelas, y tenias son tambien algunos ejemplos.
4.2 El parasito y su habitat:
Lo que hace a un parasito un parasito, es que su habitat es un organismo vivo. Esto hace que su habitat este en continua lucha contra el organimo parasitante, de hecho combatimos (en el caso de los mamiferos y aves) algunos microparasitos elevando la temperatura corporal, desarrollando reacciones inmunitarias, digiriendolo, etc, etc.
Considerando pues a los huespedes como habitas Janzen (1968,1973) introdujo la analogia de pensar en los huespedes como islas colonizadas por parasitos. De esta forma un individuo parasitado es en ultima instancia una isla colonizada, esto lleva al estudio del parasitismo desde el punto de vista de la ecologia, al mismo concepto que el estudio de la biogeografia de islas.
4.2.1 dispersion y colonizacion:
Las diferentes especies de parasitos tienen modelos de dispersion diferentes. En el caso de los microparasitos el contagio (“colonizacion”) se produce de manera proporcional al numero de encuentro que puede darse en una poblacion de posibles huespedes, por ello la propagacion de una epidemia esta relacionada con la distancia que exista entre huespedes.
El exito de la colonizacion tambien esta relacionado con la alta especificidad que suelen presentar los parasitos, ya sabemos que los organismos se encuentran muy restringidos debido a su especializacion a determinados ambientes. Como ya hemos visto en el anterior capitulo, los depredadores estan especializados en sus presas y aun siendo generalistas o polifagos la gama de presas para las que esta diseñado un predador reduce mucho el rango de estas. En los parasitos resulta si cabe aun mas intensa la relacion con su huesped, y normalmente es facil encontrar parasitos que estan restringidos a una sola especie de huespedes, y esto es asi porque el parasito esta dentro de una estrategia evolutiva estable (recordemos el apartado de la carrera de armamentos).
Por otro lado los parasitos no solo se especializan en una especie o genero, sino que ademas lo hacen sobre algun tejido especifico. Esto nos lleva a dar una nueva clasificacion:
i) Los Ectoparasitos toman su alimento desde el exterior, pulgas y garrapatas en animales y afidos en las plantas;
ii) Un grupo intermedio que desarrollaria su actividad en la cavidades internas, como son los nematodos que desarrollan su actividad en los canales alimenticios.
iii) Y por ultimo los endoparasitos, como la malaria, que desarrollan su actividad en el interior de los tejidos.
4.3 Adaptaciones al parasitismo:
La estructura general de los endoparasitos refleja normalmente las condiciones menos rigurosas, limitadas y uniformes del ambiente en el que viven. Los sistemas locomotores suelen estar reducidos, y los organos sensitivos que se encontrarian en organismos de vida libre, a veces estan reducidos o incluso han desaparecido. Y el sistema nervioso tambien se reduce a su vez.
Sin embargo los parasitos han desarrollado varias especializaciones para protegerse del hospedador. Algunos se han especializado en zonas donde el sistema inmune no llega, como es el tubo digestivo, otros poseen fases de desarrollo enquistadas y otros incluso utilizan sistemas de mimetismo (como los insectos que se parecen a hojas) con el sistema inmune y adoptan los antigenos en su superficie para no se reconocidos por el sistema inmunitario, este es el caso de los trematodos.
Un parasito con exito no mata a su hospedador, o al menos no deberia hacerlo, porque es obvio que la muerte del hospedador acarrea la suya propia. Si lo hace tendra que tener tiempo para que escape el o sus huevos.
4.3.1 Evolucion del parasitismo:
Los ectoparasitos han evolucionado a partir de especies que inicialmente eran comensales u ocupantes ocasionales de la superficie de los hospedadores. Algunos parasitos han evolucionado a partir de antecesores que estaban preadaptados a una existencia parasita. Por ejemplo, algunas moscas parasitas ponen sus huevos perforando la piel del ganado, y estas estan estrechamente relacionadas con especies que ponen sus huevos en la carne de animales muertos.
Por otra parte la boca de los hospedadores juega un importante papel en la evolucion de algunos endoparasitos. Los antecesores fueron ingeridos accidentalmente y sobrevivieron. La supervivencia podria haber originado una seleccion rapida de las carcteristicas que mejoraron las relaciones parasitarias. En aquellos que poseen hospedadores intermediarios, estos fueron quizas probablemente los hospedadores primitivos. Cuando el primer hospedador fue comido por el segundo, el parasito volvio a sobrevivir pero las adaptaciones que hicieron posible su supervivencia en el primero tuvieron que ser conservada en su ciclo biologico.
4.4 Efectos sobre la supervivencia, el crecimiento y la fecundidad.:
Una vez que el parasito o los parasitos han infestado un individuo este reducira su tasa de crecimiento y de fecundidad, asi como aumentara su grado de depredacion, de hecho tambien puede llegar disminuir la eficacia competitiva del hospededor frente a otros competidores. Un importante hecho es que puede debilitar a un competidor agresivo, y favorecer a otro no agresivo que no este infectado.
Parece que el principio “causar daño al huesped” es condicion indispensable para la definicion de parasitismo. Pero esto no siempre es tan claro y a veces el daño que se produce es mas sutil de lo que cabria esperar. Por ejemplo en la especie papamoscas cerrojillo (Ficedula hypoleuca) puede ser infectado por un Trypanosoma sanguineo, este reduce ligeramente la longitud de su cola (que es sinonimo de atrayente sexual para la hembra), de modo que cuado quieren aparearse son menos virtusos para las hembras. Pero existe un experimento en una lagartija australiana (Tiliqua rugosa), cuya longevidad no se hallaba correlacionada con la carga de garrapatas que arrastraban, y los individuos que se apareaban presentaban mayor numero de ectoparasitos que los que no se apareaban.
En general los organismos que son resistentes al parasitismo evitan los costes que produce ser parasitado, de la misma forma que sucede con la resistencia con enemigos naturales. Es decir en ausencia de parasitos una especie A susceptible al parasito sera mas competitiva que una especie B resistente al parasito. Los organismos estan sujetos a los costes de susceptibilidad y resistencia de igual forma que en el apartado 3.3.1-iii, cuando citaba el ejemplo del Guepardo.
4.5 Un caso especial, parasitismo de cria:
Hasta ahora hemos visto relaciones parasitarias donde el recurso que se explota es el huesped en si mismo robandole sus tejidos para la nutricion del parasito o aprovechando su maquinaria celular yo energia, y en casi todos los casos las relaciones filogeneticas entre ambos linajes solian estar muy alejadas, bacterias-mamiferos, artropodo-reptil, etc, etc. Pero en el caso del parasitismo de cria, las relaciones filogeneticas suelen estar muy proximas e incluso a veces se dan en el seno de la misma especie.
Se conocen ejemplos en hormigas que capturan larvas de otras especies a las que luego someten como obreras, pero son las aves las que proporcionan los ejemplos mas conocidos. Muchas hembras de gorrion domestico (Passer domesticus) son incapaces de aparearse con un macho que posea un territorio, por lo que se aparean con varios machos dominantes y depositan sus huevos en los nidos de otras hembras (parasitismo intraespecifico), con ello evitan los costos de la reproduccion delegandolos a otras parejas. Este tipo de parasitismo puede llegar a ser ocasional en numerosas especies de aves (tambien se han descrito casos en los que algunas Anatidas realizan esta actividad), pero en ciertos grupos ha pasado de ser facultativo a ser un hecho obligado. Tal es el caso que en el cuco europeo (Cuculus canorus) depende para su reproduccion de otras especies por completo (parasitismo interespecifico). Normalmente se deposita un unico huevo y en la mayoria de los casos el polluelo intenta monopolizar los cuidados de los padres adoptivos, matando (arrojando del nido) a sus “hemanastros”.
Ahora bien, este modelo de parasitismo se aproxima bastante a los modelos de parasitismo mas conveccionales, y tambien se dan procesos de carreras armamentisticas. La presencia del parasito disminuye la eficacia del hospedador, por lo que este es seleccionado para detectar su presencia y rechazarlo: bien expulsandolo del nido o bien negarse a cuidar de el. El parasito por su parte desarrolla adaptaciones para engañarlo y producir huevos y pollos mimeticos, el parasito juega con ventaja. La presion selectiva (apartado 3.3.1-i) es mas alta para el parasito ya que este se juega la vida mientras el hospedador no, si bien la de su descendencia si. Una cria de cuco puede dedicar gran parte de sus recursos hacia la adaptacion y engañar a sus padres falsos yo excluir a sus hermanastros . Sin embargo, el hospedador debe dedicar sus adaptaciones no solo ha criar mal un cuco sino a criar bien a sus hijos propios, con lo cual ambos linajes corren carreras de armamentos diferentes. Aqui suelen ganar los parasitos y estos parten con ventaja.
5. Simbiosis y mutualismo
5.1 Definicion:
La simbiosis es un termino que significa “vida en comun”, esta definicion se creo para agrupar bajo un mismo nombre a todas aquellas especies que mantienen una estrecha relacion con otros organismos de diferentes especies. A diferencia del parasitismo, las relaciones que aqui se establecen suelen ser beneficiosas para ambos actores. Pero muchos organismos se benefician unos de otros y muy discutidamente puede decirse que lleven una vida en comun (clasico ejemplo del insecto buscando polen en la flor.Mutualismo). Otras veces el beneficio va solo en una direccion aunque sin perjuicio del otro (comensalismo)
5.1.1 Mutualismo:
Se trata de una asociacion de un par de especies (generalmente), y cuyo encuentro acarrea siempre un beneficio mutuo. El mutualismo no implica una relacion fisica entre la pareja, por lo que los mutualistas no son necesariamente simbiontes. Por ejemplo; el clasico ejemplo de la abeja (Apis melifera) y la margarita. Es indudable que ambos se ven beneficiados del encuentro, la abeja obtiene alimento y la flor a cambio del polen cedido es fecundada. Sin embargo estas interacciones mutualistas no son consideradas como simbioticas. La frontera entre mutualismo y simbiosis esta poco clara pero de modo general se puede establecer que el mutualismo no implica una asociacion fisica-obligada y en la simbiosis esta asociacion se da de manera mas patente.
5.1.2 Algunos ejemplos de relaciones mutualistas:
i) El pez limpiador establece un territorio que podriamos denominar “estaciones de limpieza”. Los clientes (otros peces) los recocen y se acercan a estos puntos cuanto mayor es su grado de parasitismo. Con esta relacion mutualista los peces limpiadores obtienen alimento y el pez visitante se libra de sus parasitos.
ii) Otro ejmplo curioso es el del pajaro de la miel africano (Indicator indicator) y el tejon melifero (Melliovora capensis). Este pajaro es capaz de encontrar los escondidos panales de las abejas pero por si solo es incapaz de explotarlo como recurso, de forma que cuando localiza uno se concentra en la busqueda de un tejon y lo atrae hasta el panal. El mamifero rompe el panal y nuestro astuto pajaro se alimenta de las sobras.
iii) Los langostinos del genero Alpheus excaban madrigueras y el pez gobio Cryptocentrus se aprovecha de ellas en un ambiente donde las cavidades para la proteccion son ausentes.El langostino que es ciego mantiene contacto con el pez atraves de sus antenas y este le avisa en caso de peligro. Las ventajas que ambos obtienen son claras.
iv) Los mutalismos de dispersion de semillas son el resultado de una larga coevolucion entre plantas aves y mamiferos. Los beneficios de que la planta obtiene son unicamente la dispersion de semillas, pero no es poco ya que con ello las plantas se alejan aleatoriamente de su area “materna” y colonizan nuevos territorios, por otro lado los animales que la consumen obtienen alimentos ricos en azucares y lipidos. La costosa inversion que tiene que hacer las plantas esta en el fruto, que se ve recompensado por la dispersion de las semillas.
5.1.3 Mutualismos que implican cuidado de ganado y cultivos:
Las hormigas son un grupo especializado en este tipo de mutualismos, en areas del Amazonas existen generos como Atta y Acromyrmex que se dedican a recolectar hojas de arboles y asi cultivar en ellas hongos del genero Hypomyces, el hongo es cultivado para el consumo pero con la hormiga su supervivencia esta asegurada. Otras hormigas mas conocidas como las de jardin (genero Lasius) establecen una actividad mutualista con insectos hemipteros. Los pulgones parasitan las yemas de las plantas, y las hormigas como ya sabemos precisan de un aporte de azucares en su dieta ( que dicho sea de paso es simpre omnivora), precisamente estos “animalillos” pueden proporcionarles toda esa “azucar” que necesitan. Los pulgones chupan mas de lo que necesitan puesto que lo que ellos buscan realmente es nitrogeno, el subproducto es expulsado por la parte posterior superior del abdomen y las hormigas solo tienen que recogerlo. Este relacion otorga a las hormigas “comida” y a los pulgones una estimable defensa contra cualquier prededor.
5.1.4 Otros tipos de mutualismos:
Como podemos ver existen relaciones mutualistas a la carta, intentar por tanto sintetizar todos en un trabajo como este seria tarea imposible y cuanto menos redundante. Por ello y al margen de los que merecen la pena ser mencionados (como el rumen ), citare algunos de las relaciones mutualista de mayor importancia:
El rumen: La mayoria de los intestinos de los animales es un microcosmos de vida bacteriana y protozoos. En ellos se da unas condiciones de Ph y temperatura mas o menos constantes y se suelen dar condiciones de anaerobiosis. Los mamiferos pequeños tienen asignado una parte de sus intestino para la fermentacion (el ciego) de los vegetales que comen y en animales grandes como el caballo el ciego es el lugar mas importante para la fermentacion.
En los animales herbivoros rumiantes (ciervos, vacuno, antilopes, etc.) la camara fermentativa se encuentra a nivel del estomago, esta dividido en 4 compertimentos. La primera masticacion conduce el alimento a la camara del rumen de ahi pasa al reticulo a medida que su tamaño va disminuyendo, de ahi pasa a la tercera camara, el omaso, una vez ahi el animal regurguita y se produce una segunda masticacion que reduce aun mas las particulas vegetales. Lo verdaderamente curioso es la actividad microbiana que se da en el rumen, y los productos de la fermentacion que ellos generan es lo que alimenta al huesped. A su vez es el huesped el que genera las condiciones necesarias para este proceso. Estas bacterias y protozoos son anaerobias estrictas y si entran en contacto con el oxigeno mueren. La mayoria de las especies que forman el rumen poseen tal especializacion que no podrian vivir fuera de el.
La cuestion de esta relacion es la carencia por parte del rumiante del aparato enzimatico para degradar la celulosa y otras fibras indigeribles en la dieta de los rumiantes. Bacteroides succinogenes digiere solo celulosa, Ruminococcus celobiosa y celulosa, otros pueden digerir tambien el almidon como es el caso de Clostridium locheadii. Ahora bien, como ya dije antes el tracto digestivo de los animales puede ser un microcosmos de vida, por tanto la flora del rumen esta sujeta a fenomenos de competencia en una escala inferior, al tratarse ademas de un ambiente constante (como ocurre en los tropicos), los fenomenos de competencia lejos de darse deben ser muy intensos.
Los liquenes: Numerosos hongos han escapado a su habitual sistema de vida libre y se han asociado con algas. Estos hongos son los denominados hongos “liquenizados”, contienen una capa de celulas fotosinteticas cerca de la superficie (algas).
Los hongos obtienen del alga productos de la fotosintesis y en algunos casos nitrogeno. La ventaja para el alga no esta muy clara y aun es objeto de investigacion, solo en algunos generos como Trebouxia parece que debe existir alguna relacion de beneficio porque las formas de vida libre son raras.
No es asi en el caso del hongo, las formas liquenizadas son diferentes en morfologia de las formas libres por lo que el crecimiento de este se halla profundamente alterado por la presencia del alga, son ademas eficientes colonizadores y su dispersion como colonizantes es mayor cuando estan asociados.
5.2 Fijacion del nitrogeno y su relacion con el mutualismo:
Como ya sabemos el nitrogeno es fundamental para la vida y es el elemento constituyente de las proteinas, en la naturaleza lo encontramos formando parte de los seres vivos, de forma inorganica como amonio, nitrito y nitrato (formas asimilables por las plantas), y en la atmosfera donde esta en forma de N2. El nitrogeno atmosferico representa casi el 80% de todo el nitrogeno del planeta por lo que su disponibilidad para los seres vivos hace que sea uno de los elementos mas limitantes ( la mayoria de los organismos no pueden asimilarlo en su estado gaseoso). Sin embargo existen organismos capaces de asimilar el nitrogeno atmosferico.
Azotobacter, Azotococus, Rizobios, Clostridiun spp, Enterobacter y/o cianoficeas en general son algunos ejemplos de bacterias que sintetizan el nitrogeno del aire hacia formas asimilables por otros organismos (plantas). Pero la familia Rhizobiaceae es de todos los grupos la mas estudiada.
El contacto entre el rizobio y la planta huesped (siempre una leguminosa) se produce a nivel de los pelos radiculares de la raiz, la bacteria que se encuentra en estado de vida libre es estimulada para la multiplicacion por los exudados de la raiz. Estos exudados son los responsables de la activacion de una serie de genes de la bacteria, que controlan tambien el proceso de nodulacion. De forma general se desarrolla una colonia alrededor de un pelo radical este se riza y forma un tubo de infeccion por donde pasan las bacterias. Una vez estas han penetrado en las celulas del huesped, se produce la formacion del nodulo.
El huesped entonces desarrolla un sistema vascular “especial” que retira del nodulo los produtos de nitrogeno fijado y aporta nutrientes fotosinteticos. Dentro de los nodulos existe una proteina homologa a la hemoglobina, la leghemoglobina, que posee una alta afinidad por el oxigeno, favorciendo asi que el nodulo se mantenga anaerobico (hay que recordar que el equipo enzimatico de la fijacion de N2 se realiza en anaerobiois) para la nitrogenasa.
5.2.1 Efectos del mutualismo fijador de N2 atmosferico en los ecosistemas:
La fijacion de nitrogeno se reduce en los ambientes que son ricos en N2, pero si eventualmente excasea la fijacion se dispara. Esto conlleva que todas las especies vegetales con asociaciones mutualista obtengan un beneficio neto superior al resto de las plantas. Por lo que en los ambientes con deficiencia de nitrogeno, las leguminosas fijadoras llevan ventaja frente al resto de espescies. Sin embargo existe una paradoja en este hecho, y es que a medida que el ambiente es enriquecido con Nitrogeno, las otras especies vegetales tambien se benefician, por lo que la ventaja inicial se pierde ya que el ambiente ha sido mejorado para los competidores . Quizas esta sea una de las causas de porque las leguminosas (tanto herbaceas como arbustivas) no hayan dado lugar nunca a poblaciones dominantes en la naturaleza.
De cualquier forma la importancia de los organismos fijadores de nitrogenos (a parte de las relaciones mutualistas aqui descritas) es incalculable. Los aportes de nitrogeno que realizan de manera natural es la unica via de entrada de nitrogeno atmosferico al ecosistema.
5.3 Eucariotas, producto de una simbiosis:
Hemos podido ver hasta ahora un compendio de ejemplos que nos hace ver como organismos diferenten se hacen “socios” y en algunos casos pueden llegar ha convertirse en una especie de organismo fusionado. Pues aplicando este concepto Margulis (1975) desarrollo una atrevida hipotesis (hoy teoria endosimbiotica) sobre la evolucion de la celula eucariota.
Segun Margulis, la evolucion de los eucariotas debio producirse por interacciones de procaritas con otras procariotas, es decir quizas un dia una bacteria con el ciclo de Krebs fue incluida dentro de otra, sobreviviendo una dentro de la otra. Mas tarde se asociaron a otras que poseian cilios o flagelos y asi hasta que despego la evolucion de estos primeros procariotas. Esto explicaria el origen de los organulos celulares, cloroplastos y mitocondrias poseen ADN y gozan de cierta autonomia con respecto al resto de la celula.
6. Competencia Intraespecifica
6.1 Definicion, efectos y caracteristicas de la competancia intraespecifica:
Ya hemos visto que los organismos se comen, se engañan y se asocian, pero lo que aun no hemos visto es que tambien son victimas de sus propias actividades. No existe ningun organismo que viva aislado. Y al menos una vez en su vida tendra que reproducirse y siempre formara parte de una poblacion con los de su misma especie.
Las necesidades que un organismo posee son invariables con las del resto de individuos de su misma especie, esto unido a la necesidad de no perecer de inanicion o espacio para criar, puede acelerar la demanda de un determinado recurso y agotar la oferta del mismo. Entonces el recurso se vuelve raro y los organismos compiten por el.
Consideremos el siguiente ejemplo: una poblacion saltamontes que crece y vive en un campo de herbaceas. Para crecer y reproducirse los saltamontes necesitan 2 cosas materia y energia; pero para obtener esto tambien precisan energia para desplazarse. Si un saltamontes llega a una parcela donde no hay comida tendra que volar para encontrar otra parcela donde no halla llegado un saltamontes antes que el. Por ello la supervivencia del saltamontes estara condicionada a la capacidad de encontrar antes de que otro lo haga ya que en ello va su eficacia reproductora o su propia supervivencia.
Pero en realidad esta no es una competencia directa entre individuos, el saltamonte ni se percata de que hay o ha habido otros congeres en su parcela, solo responde al hecho de que no hay comida y la necesita. En otros ejemplos si puede darse una competencia directa entre individuos (Interferencia), el individuo que accede primero al recurso impedira su acceso a otros congeneres, Por ejemplo entre Septiembre y Octubre se registra en la peninsula Iberica el fenomeno de la berrea, los machos del ciervo (Cervus elaphus) luchan entre si para defender un haren de hembras y un territorio, aqui el recurso esta claro; hembras para la reproduccion, y este es un un recurso limitado.
De este modo la competencia por interferncia puede desarrollarse por un valor real (comida, espacio, hembras, etc) o por sustituto (la conquista de cierto territorio implica la posesion de un haren).
6.2 Natalidad y mortalidad:
De manera general se puede decir que la mortalidad de una poblacion en situacion de estres por una alta densidad, puede ser dependiente de la densidad, esto significa que la mortalidad se incrementa con la densidad. Esto es facil de entender si volvemos al ejemplo anterior del saltamontes. Si partimos de una poblacion X de saltamontes que crecen y se reproducen y controlamos su tasa de mortalidad, veremos que a medida que aumenta su densidad aumentara su mortalidad, y esto ocurre por hecho muy sencillo; a medida que crece la poblacion disminuye la tasa per capita de alimento para cada individuo, los fenomenos de competencia por el alimento aumentan, y solo sobreviven los individuos mas competitivos.
En el caso de la natalidad, los resultados obtenidos por los investigadores son casi una imagen especular de la mortalidad, por lo que los efectos de la competencia intraespecifica ocasiona de igual modo un descenso de la natalidad, y a largo plazo contribuye al mismo efecto sobre la poblacion que la mortalidad; la poblacion decrece.
Para este tipo de estudios sobre el crecimiento de poblaciones animales existen numerosos modelos matematicos que intentan explicar como evolucionan las poblaciones sujetas a fenomenos de competencia. Para poblaciones con estaciones de reproduccion que pueden escribirse mediante ecuaciones en “diferencias” (aumentan en pasos discretos), existen modelos que analizan como varia de tamaño la poblacion.
6.3 Ecuaciones en diferencias:
Estas ecuaciones nos sirven para teorizar como se comportara una poblacion bajo fenomenos de competencia intraespecifica. El modelo consta de dos ecuaciones:
t
Nt+1 = NtR y Nt= N0R
Nt representa el tamaño de la poblacion en el momento t, y R representa la tasa de reproduccion bajo la cual la poblacion crece.
Pero este modelo por si solo no nos resuelve nada, en el se describe una poblacion carente de competencia, por lo que si R$1, la poblacion creceria de manera exponencial ( esto es lo que ocurriria en la naturaleza con alimento y espacio “ad libitum”). Modificando ligeramente las ecuaciones y teniendo en cuenta los factores de competencia sobre la tasa de reproduccion, podemos obtener una situacion donde Nt sea un valor muy reducido (casi cero) y por tanto la competencia despreciable (recordar que los efectos de la competencia son patentes a altas densidades de poblacion), obteniendo:
Nt/ Nt+1= 1/R La tasa de reproduccion no queda altarada.
Pero a medida que Nt aumenta, la tasa de reproduccion se altera de manera que cuando el cociente Nt/ Nt+1 es igual a “0” la poblacion deja de crecer y solamente se sustituye en cada generacion. El tamaño de la poblacion por el que se produce este fenomeno es denominado capacidad de carga, K.
Si tenemos en cuenta la grafica A y B son los puntos que describen la tasa neta de reproduccion a medida que lo crece la poblacion. Nt/ Nt+1 se mide en el eje y, Nt en el eje x, y la interseccion es 1/R, la pendiente es por tanto
Nt/ Nt+1= 1-1/R/K x Nt + 1/R
Reorganizando:
Nt+1= Nt R/1+(R-1) Nt/K simplificando, (R-1) con una “a”:
Nt+1= Nt R/(1+aNt)
Este es el modelo definitivo, donde R a pasado de ser una constante a ser la tasa real de reproduccion, R/(1+aNt), que disminuye a medida que aumenta el tamaño de la poblacion Nt.
6.4 La ecuacion logistica:
Ya hemos visto muy a grosso modo el modelo matematico con ecuaciones continuas, y como este nos sirve para hacer estudios sobre el comportamiento de las poblaciones. Sin embargo para poblaciones donde las muertes y los nacimientos se producen de manera continuada, se utilizan modelos donde las ecuaciones mas apropiadas son las “logisticas”.
Aqui el crecimiento de una poblacion se denotara como dN/dt. Esto nos representa la velocidad de crecimiento N, en un tiempo t. Asi pues la tasa de incremento per capita es dN/dt(1/N), pero como esta expresion no recoge la competencia, suponemos que lo que medimos es en ausencia de competencia:
dN/dt=rN
En esta expresion donde no existte competencia, la poblacion crecera exponencialmente como se ve en la grafica. Para ajustar el modelo a la realidad debemos introducir la competencia intraespecifica. Si miramos la siguente grafica observamos que la tasa neta de incremento por individuo no se ve afectada por la competencia si N esta cerca de “0”. Cuando N aumenta hasta K, la tasa neta por individuo es cero. Teniendo en cuenta la linea A, B :
dN/dt(1/N)= (-r/K x N)=r que simplificando:
dN/dt(1/N)= r(1-N/K)
dN/dt= rN(K-N/K)
Estas ecuaciones han desarrollado un papel fundamental en el desarrollo de la ecologia. Es importante destacar tambien que se tratan de modelos muy limitados, pero he considerado sin profundizar en ello al menos representarlas.
7. Competencia interespecifica
7.1 definicion:
Al igual que ocurria con la competencia intraespecifica, se produce una disminucion de la tasa de natalidad, aumento de la mortalidad y una disminucion del indice de superivencia individual, debido a que los recursos que explota una determinada especie son tambien explotados por otras especies diferentes y a veces con mayor efectividad. Este hecho lleva a las especies a evitar explotar el mismo recurso para no solaparse y que la coexistencia sea posible. Sin embargo tambien se dan ejemplos en la naturaleza de exclusion de una especie por competencia si el recurso es lo suficientemente limitante.
7.2 Principio de la exclusion competitiva
Este principio puede resumirse en que dos especies no pueden coexistir indefinidamente sobre la base del mismo recurso limitante (ley del minimo de Liebig). Por lo que este principio predice que dos especies con iguales requerimientos no pueden existir. Sin embargo en la naturaleza se dan este tipo de coexistencias, pero como vamos a ver a continuacion esto es posible debido a que las diferencias aun siendo sutiles se dan.
7.3 Competencia en espacios desocupados
Cuando una especie excluye a otra es porque compiten en terminos de igualdad, la coexistencia es posible cuando cada especie se apropia de espacios diferentes (aunque tambien hemos visto que no es totalmente cierto). Esto es lo que puede ocurrir cuando en un ecosistema se produce una “perturbacion”. Tras una perturbacion (un incendio, un desprendimiento de tierra, etc.) el exito de los organismos que alli se instalen dependera en gran medida de lo vacantes que esten los nichos ecologicos que ocupen. Llegar primero tambien puede inclinar la balanza a la primera especie que lo consiga.
7.4 Competencia entre paridos:
El carbonero comun (Parus major), herrerillo (P. caeruleus), el carbonero palustre, el sibilino y el garrapinos (P. palustris, P. montanus y P. ater, respectivamente), son especies que pueden aparecer todas juntas en bosques de pinos y montaña. Todas estas aves nidifican en agujeros, todas comen insectos durante todo el año, todas buscan las mismas semillas durante el invierno y sin embargo, coexisten. Esto era posible porque siendo tan parecidas las especies de esta familia, habia algo que lo permitia, y es que cada ave estaba especializada en un estrato arboreo diferente, en un tamaño de insecto diferente y al parecer en una dureza de semilla diferente. Gracias a esto las especies coexisten porque explotan recursos ligeramente diferentes.
7.5 Competencia con diatomeas:
Este otro ejemplo ocurre justo lo contrario. Se trabajo con dos especies de diatomea de agua dulce: Asterionella formosa y Synedra ulna (Tilman et al.,1981). Como ya sabemos las diatomeas son algas unicelulares que requieren de silicatos para poder construir sus paredes celulares. Cultivando ambas especies por separado se observo que ambas poblaciones se mantuvieron estables. Sin embargo, Synedra consumia mas silicato que Asterionella, por lo que cuando ambas especies convivian Synedra dejaba los niveles de silicatos demasiado bajos como para que Asterionella pudiera sobrevivir.
7.6 Modelo matematico pera la competencia interespecifica:
Un modelo matematico que intente explicar como varian poblaciones con competencia interespecifica debe tener en cuenta que por lo que se compite es por los recursos, para ello existe un modelo logistico. El modelo de “Lotka y Volterra”
dN/dt= rN(K-N/K)
Ya hemos visto con anterioridad como este modelo explica los fenomenos de interacciones entre especies. En este caso el tamaño de la poblacion viene indicado por N1 y el de la segunda especie por N2. Sus capacidades de carga y sus tasas intrinsecas vienen indicadas por K1, y r1 respectivamente.
Para entender este modelo en la competencia interespecifica vamos a suponer que se dan dos especies A y B, de forma que al menos 10 individuos de la especie B ejerce el mismo grado de competencia que 1 individuo de la especie A. El efecto total que actua sobre la especie A sera equivalente al efecto de (N1+N2/10) individuos de la especie A. La constante 1/10 se llama coeficiente de competencia y se simboliza por α12.
De esta forma con estas dos expresiones se trabaja para el analisis de las especies A y B:
dN1/dt= r1N1(K1-N1- α12N2)/K1
Y en la segunda especie:
dN2/dt= r2N2(K2-N2- α21N2)/K1
Pero todo esto es mentira! las especies no evolucionan ni coevolucionan ni se adaptan ni interaccionas segun algunos...
Fuente BIBLIOGRAFIA Y REFERENCIAS
· Lovelock, Margulis, L. Bateson, G. Maturana, H. Varela, F. et all. (1989). “A way of knowing”. Edt. Kairós, S.A. Edt. The Lindisfarne Association
· M. Begon, J.L Harper, C.R.Towsend. (1999). “Ecology: Individuals, populations and comunities”. Tercera edición. Editorial Omega, S.A., Barcelona.
· Wilhelm Goestsch. (1987). “Die staaten der Ameisen”, La vida social de las hormigas. Segunda edición. Editorial Labor, S.A., Calabria.
· Edward E. Rupert & Robert D. Barnes. (1996). “Zoología de los Invertebrados”. Sexta edición. Editorial McGraw Hill Interamericana editores, S.A, México
· Universidad de Extremadura. (1994). “ETOLOGIA, introducción a las ciencias del comportamiento”. Tercera edición. Editorial Servicios de publicaciones de la universidad de Extremadura. Cáceres.
zarasein05 de jun. 2007Apomios a mí también me gusta mucho leer El Corán, y mucho más que la actual biblia judeo cristiana, ya que al menos, contiene algunos pasajes bien traducidos de la BIBELE "el Destino de los Bele" original .
Mas hay convicciones y creencias personales que es mejor no airearlas a la ligera en público ya que muchas de ellas forman parte de los más íntimo y personal de cada uno de nosotros y si las exponemos en lugares y ante gente inadecuada corremos el riesgo de ser heridos en lo más profundo de nuestro ser.
Si a un ateo materialista educado en la teorias neoevolucinistas le explicas que la palabra DIOS significa nada más y nada menos que "El Inteligente" o que también se podría traducir por "Nuestra Suprema Inteligencia" y que la pronunciación correcta sería ADIO, muy parecido al "adios" castellano o al "adeu" catalán, lo más probable es que me tome por chiflado. Si a continuación le explico que ese Ser Inteligente al que me refiero también lo llamamos JAUNGOIKO: "Señor de los Cielos" y que se denominaba así a Aquel de entre los más sabios del Cielo, quien había conseguido desentrañar los misterios de Râ siendo capaz de crear como él y perfeccionar sus creaciones y que para los Bere, Jaungoiko era el Gobernante de los Cielos al cual debían obediencia en honor a su demostrada sabiduría y justeza, pues probablemente intentarán ponerme la camisa de fuerza y en el mejor de los casos cerrarme la boca.
Pero como tengo más que comprobado que Dios cierra los ojos y los oidos a quien El quiere, pues ya no gasto inútilmente mis energias en debatir ni discutir nada con esa gran masa de humanos perfectamente aleccionados para que rindan culto y veneración solo al dios dinero, o a políticos mentirosos y corruptos que les prometen un mundo mejor gracias a ellos y que consuman compulsivamente cosas innecesarias hasta endeudarse hasta los ojos con los Bancos y Cajas que juntos con los grandes trust energeticos son los verdaderos amos de este mundo donde a Dios, según ellos (qué poco inteligentes que son), lo mandaron a la guillotina con la revolución francesa.
Lo han montado tan bien su "tinglado", que a los creyentes nos consideran poco menos que fanáticos acientíficos que no merecen respeto ni consideración alguna. Mas lo que ellos no saben es que TODAS LAS CIENCIAS CONDUCEN A DIOS Y QUE SOLO DIOS REVELA TODAS LAS CIENCIAS.
He extraído de la web de TARTESSOS un artículo sobre la BIBELE original que puede ilustrar muy bien lo que quiero decir sobre Dios(Allah, Jaungoiko o como le queramos denominar) , que al ser el más Sabio, pues sin duda es el mejor cientítico de todos los humanos y que su CIENCIA no tiene fin.
"En el Bibele el Zar de los Ele explica en síntesis la Historia de la Humanidad desde sus orígenes hasta el Día en que será pronunciado su Juicio Final, la razón de su Misión, las incidencias de su viaje interestelar rumbo a la Tierra y también el Sistema empleado para lograr su Primer Resurgimiento.
Escrito está en la Bibele original: Te envío del Cielo el Libro que contiene la Verdad de tu destino y de las tres humanidades -la blanca, la negra y la amarilla- para que puedas juzgar a los hombres según el modelo nuestro. No te enfurezcas con los impostores. Serán ridículos y se mofarán de ti pero tú conoces la Verdad del futuro y ellos no. Conocerás cada vez la Gloria de tus dos nuevos Resurgimientos y ellos la espantosa miseria del suplicio del fuego eterno.
Es cierto que Dios Todopoderoso envió del Cielo el Libro esclarecedor. Envió el Aerolito, la KAIXE, en la cual estaban grabadas las instrucciones de Dios dirigidas a Iezus o Goberiel, referentes a las Tres Misiones que EL debía cumplir en el transcurso de sus Existencias en este Globo. Misiones que debían acontecer según un plan y unos tiempos preestablecidos hasta el Día del Juicio.
El Aerolito cayó en un lugar que a partir de entonces sería llamado ZERUAMEKA "El caído del Cielo", siendo hoy La Meca un lugar desde entonces Sagrado y Venerado. Iezus en el Libro también revela detalladamente nombres, lugares y fechas así como las razones de Justicia y Conciencia por las cuales hará su aparición en dos próximas vidas en esta Tierra, antes del Día Fatídico. En el Bibele dejó también escrito qué es lo que seguiría después de su Juicio, que es un final con dos alternativas: o bien la Redención de toda o parte de esta Humanidad o el Golpe Final que está programado a un Día Tope e inevitable. La Bibele, por tanto, es una Obra histórica, geográfica y científica de inmenso valor debiendo considerarse como el Libro más importante escrito en esta Tierra. Es la herencia de Iezus-Eleazar a los Bere quienes pueden ser considerados como la verdadera Gente del Libro.
EL escribió: La obra escrita del primer libro tuyo servirá de prueba y testimonio de la alta capacidad y justicia de tu ciencia.
El actual libro -hoy muy famoso- conocido como La Biblia de los católicos, no tiene nada -o casi nada- que ver con la genuina de la cual sólo se conserva su título y algunas frases muy amañadas, resumidas y deformadas en su primer capítulo: el Génesis. El resto es una patraña, mezclada con ciertas leyendas muy retocadas por los sacerdotes paganos los cuales no han tenido ningún reparo en reclamar a cada página de la misma se les den tributo en carneros o monedas de oro y plata, cada vez que alguien respira o emprende alguna labor. Como no pudieron acabar con la gran fama y arraigo popular del Libro, idearon crear otro que llevaría su mismo título pero cuyo contenido sería el por ellos convenido, más acorde a sus intereses mentirosos. Ese libro es la Biblia que todos ahora conocemos y puede considerarse como una auténtica blasfemia a Dios Todopoderoso y a Dios Be.
Hoy día, apenas quedan algunos ejemplares de la Bibele original repartidos en diversos países y custodiados por sus celosos guardianes. Su sola posesión es peligrosa siendo motivo de persecución y de muerte. La mayoría han sido destruidos a conciencia por los Pares, los sacerdotes paganos y los idólatras de todos los tiempos.
Escrito está en el Libro que esclarece el entendimiento: Zar de los Ele, verás como los profesores de la cultura prostímera de los idólatras corromperán esas escrituras nuestras. Esconderán los textos auténticos que hablarán de la Ciencia y del Poder futuro. No deberás parar un instante en descubrir sus fechorías, sus falacias, sus fraudes.
El manuscrito original de la Bibele debió permanecer algún tiempo en la tumba de Iezus, siendo después expuesto al público en Bibelos y Balbek. Mucho más tarde en el tiempo fue guardado en la floreciente ciudad de Damasco. De dicha ciudad sería retirado con toda urgencia al ser atacada por los Panie de Palemere (Palmira) y los Palestinos, los cuales destruyeron la Mezkite con la tumba Sagrada de la Mere (Maria). Así retirada, la Bibele fue trasladada a La Meca, donde se la comenzó a llamar también ELEBERU. Allí permaneció hasta la Victoria de Troia.
Mahoma, al escribir su libro conocido como El Corán, se refiere bastantes veces al texto original en sus versículos. Cita algunos pasajes de la Verdadera y Primitiva Bibele, tal como fueron escritos y en buena traducción. Pero, también alude a otros los cuales no supo interpretar, dejándolos a medio explicar o confundiéndose totalmente. No cabe duda de que Mahoma encontró en su gruta, cerca de Medina, una copia de la Bibele en escritura seguramente siríaca o cúfica escrita en la antigua lengua árabe. Pero, tal y como se explica en el Corán, es de suponer que tan sólo hallase una copia abreviada de la Bibele, o algunas hojas sueltas constituyendo el principio de una transcRIPción original. Además, posteriormente a Mahoma, el Corán fue retocado en algunos versículos así como se amañaron otros. Pero, con todo, posee auténticas partes de la Bibele original, al contrario de la Biblia utilizada por los católicos.
http://www.tartessos.info/html2/bere01.htm
1) La teoría no puede explicar por ningún medio cómo se originó la vida sobre la tierra.
Falso
La Teoría de la devolución no tiene por objeto solucionar el origen de la vida, sino la ingente variedad de seres vivos que hay sobre la tierra y la relación que existe entre los seres vivos.
2) No existen hallazgos científicos que muestren que los “mecanismos evolutivos” propuestos por la teoría tengan algún poder para provocar la evolución
Falso
Para lo que desde luego no existe es para el Creacionismo.Ya que el paradigma de la creación “una vez y para siempre” establecido como un orden inamobible por la ciencia esta tan estrechamente vinculado a las creencias religiosas y aun mas ala idea de un orden político estable,( y ya vemos que 200 años después también) que dudar sigue siendo acto cuanto menos subversivo.
Superar esto era una tarea destinada destinada a algún héroe prometeico, un rebelde con vocación. El hombre llamado a hacerlo fue Charles Darwin, un vicario rural de temperamento prudente que había decidido pasar cinco años de su vida a bordo del Beagle, un barco de la armada Británica con la misión de estudiar la fauna y flora de sudamerica.
PRUEBAS y EVIDENCIAS
En Argentina y en Chile Darwin camino entre restos de mamíferos recientemente extintos y se pregunto a si mismo, por qué eran tan similares a las formas vivas, si no fueran de algún modo sus antepasados.
En ccGalapagos, una Isla de origen volcánico de menos de 2 mill de años, Darwin se encuentra con algo mas de 18 especies de pinzones, todos diferentes y que explotaban recursos diferentes, mientras que en las costas de sudamerica aparece solo una.
Por fin en 1859, Darwin publica “sobre el origen de las especies mediante métodos de selección natural. Lo bueno de esto es que por fin su libro pudo de alguna manera ofrecer un marco en le que la ciencia podía organizar una masa de información hasta entonces confusa o demasiado confusa para tener un significado. Fue entonces Darwin quien proporciona la base estructural, el primer mecanismo convincente del cambio a largo plazo.
Los organismos producen generalmente mas descendencia de la que puede sobrevivir y reproducirse.
Los descendientes que sobreviven tienden a ser los mas fuertes.
Los caracteres de los padres aparecen en los hijos
De este modo, generación tras generación, a través de cientos de miles de años y/o periodos, las lineas mas fuertes sobreviviran al traspasar los rasgos que los han hecho fuertes.
a. Los restos fósiles prueban exactamente lo contrario de lo que sugiere la teoría de la evolución.
Falso
La vida ha cambiado a través del tiempo. No hay otra expoliación posible para la secuencia y variedad de formas de vida preservadas como fosiles o para la historia desde que el joven ser humano comenzo a dibujar, pintar y grabar, hace aproximadamente 30.000 años. Ninguna otra cosa podría explicar las relaciones entre las especies vivientes reveladas por las técnica de la moderna Biología. No solo han cambiado han evolucionado!. Toda la vida en la tierra esta interrelacionada, en un modelo que se remonta a las primeras formas de vida que aparecieron en la Tierra hace unos 4.000 mill de años. Cada planta cada animal, todo en los 5 grandes Reinos, vivo o extinto, puede ser expuesto- si pudiéramos seguirle la pista a todo- como un sistema unico de sendas, caminos muchos, recodos, quiebros, pero todoconduce a un anterior y único sendero, una especie que fue el ancestro común de todos nosotros: mohos, bacterias, maRIPosas y “ algunos capullos”...., ostras y cazadores de ostras, plancton y personas.
LA PRIMERA ETAPA INSUPERABLE: EL ORIGEN DE LA VIDA
La teoría de la evolución postula que todas las especies vivas evolucionaron de una única célula que surgió en la tierra primitiva hace 3.800 millones de años. Cómo una única célula pudo generar millones de complejas especies vivas y, si tal evolución realmente ocurrió, porqué no pueden observarse vestigios de ella en los restos fósiles
Falso
Existe toda una gama de microfosiles bacterianos ( arquebacterias, cianobacterias), en rocas que datan desde el paleozoico (3.800 mill de años), que demuestran una gradación de complejidad.
Hoy, terminando el siglo XX, enfrentamos todavía el mayor problema sin resolver que teníamos cuando comenzó el siglo: ¿cómo se originó la vida sobre la tierra?” (48).
Verdadero, pero el tema que abordamos no es el origen de la vida sino del hecho evolutivo, en cualquier caso, ya existen hipótesis que no teorías mas cercanas a la realidad que el cuento de Adan y la costilla.
Charles Darwin vio un camino por el que pudo haber maniobrado la evolucion, pero no fue el primero en reconocer que la vida había cambiadoy estaba aun cambiando. Huesos fosiles demasiado grandes para pertenecer a especies vivientes, descubiertos al arar o en excavaciones en el pasado predarwiniano, fueron identificados como pertenecientes a monstruos, gigantes levanta piedras, y Dragones que escupian fuego.
Los estudiosos de la época ( y alos iluminados de hoy en dia) explicaban que la vida debía de haber comenzado con un vagón lleno de criaturas ( y entiendase como una metáfora...),y tal vez, ellas aun estaban al acecho en las fronteras de nuestro mundo conocido. El brillante naturalista Francés ( que no Biólogo como antes se ha dicho), Jean-Baptiste Lamarck, que introdujo palabras claves como “biología”, e “invertebrado”, fue uno de los muchos pensadores del S XVIII que creía en una “escala naturae”, la escalera de la naturaleza. El visualizo una cadena, o cadenas, del ser- posiblemente para animales y otra para plantas- donde cada estadio iba de los animales mas simples a los mas complejos,y cada uno de ellos susceptible de transformarse en la siguiente a los largo de esa linea.
EL IMPACTO DE LAMARCK
Ahora bien, ¿cómo pueden ocurrir estas “variaciones favorables”? Darwin trató de responder a esta cuestión partiendo del punto de vista que el primitivo estado del conocimiento científico poseía en su época. Según el biólogo francés Lamarck, que vivió antes de Darwin, las criaturas vivas transmitían los rasgos que adquirían durante su vida a la siguiente generación, y estos rasgos acumulándose de una generación a otra provocaban la aparición de nuevas especies. Por ejemplo, según Lamarck, las jirafas evolucionaron de los antílopes; esforzándose por comer hojas de árboles altos sus cuellos fueron estirándose de una generación a otra.
Darwin da también ejemplos similares en su libro “El origen de las especies”. Por ejemplo, dice que algunos osos que se introducían en el agua para buscar comida se transformaron en ballenas con el paso del tiempo (51).
Pero no obstante, las leyes de la herencia descubiertas por Mendel y verificadas por la genética que floreció en el siglo XX, finalmente destruyeron la leyenda de que los rasgos adquiridos se trasmitían a las generaciones subsiguientes. De esta forma la selección natural perdió sustento como mecanismo evolutivo
Falso
Lamarck promulgo el primer concepto de cambio y evolucion, pero a través de un mecanismo erroneo. Darwin mejoro notablemente la idea con otro mecanismo que nada tiene que ver con Lamarck.
Los organismos producen generalmente mas descendencia de la que puede sobrevivir y reproducirse.
Los descendientes que sobreviven tienden a ser los mas fuertes.
Los caracteres de los padres aparecen en los hijos
De este modo, generación tras generación, a través de cientos de miles de años y/o periodos, las lineas mas fuertes sobreviviran al traspasar los rasgos que los han hecho fuertes.
Lo que la genetica mendeliana hace es consolidar las tesis de Darwin e invalidar a Lamarck, pero en ningún caso desacredita a Darwin.
EL NEODARWINISMO Y LAS MUTACIONES
Sin embargo, existe un hecho científico innegable que socava esta teoría: las mutaciones no provocan una evolución en los seres vivientes; por el contrario siempre los perjudican.
Falso otra vez!
Las mutaciones en su mayoría suelen ser neutras, estas pueden ser por deleccion, translocacion o conversión, y existe una mutación por la deleccion de una sola base nitrogenada en el gen relacionado con la hemoglobina que produce anemia falciforme, una enfermedad genética, sin embargo existe un una zona en el mundo donde la anemia falciforme no es dañina y no es un handicap para los que la portan, sino todo lo contrario ya que en determinados ambientes donde se da la malaria los portadores son resistentes a la malaria. Ya ves como el medio esta selecionando un caracter a través de una mutación!!!!!
RESTOS FÓSILES: NO HAY RASTROS DE FORMAS INTERMEDIAS
Falso
Ya se ha dicho que si, y sino pasate por el museo de Historia natural de Madrid, y si me dejas un consejo te recomiendo especialmente el de Marsella Francia donde hay expuesto una colección de fosiles que desarrollan todas las formas intermedias en la evolucion del Caballo./
LAS ESPERANZAS DE DARWIN SE HICIERON PEDAZOS
Un paleontólogo británico, Derek V. Ager, admite este hecho aunque él es un evolucionista:
“La cuestión que surge es que, si nosotros examinamos en detalle los restos fósiles, sea a nivel de órdenes o de especies, encontramos —una y otra vez— no una evolución gradual, sino una explosión repentina de un grupo a expensas de otro” (54).
Bueno quien quiera que ha escrito esto es que no se entera, mezclas todo hijo mio...
Derek V. Ager, es ciertamente un paleontologo evolucionista, pero al igual que Stephen jay Gould, ambos han mejorado la Teoria de la evolucion con un nuevo concepto que es el “equilibrio puntuado”
Leete “el primer ministro y la nalga del brontosaurios Stephen Jay Gould” y puede que ...bueno dejalo.
Y así se puede seguir eternamente porque lo que no puedes pretender es que creamos en Gigantes que levantan piedras, Dragones, boxeadores que son Neanderthales, y toda la sarta de sandeces que propones.
Se te dan pruebas , evidencias, hechos, y tu niegas la verdad y en contraposicion ofreces versículos coránicos de dudosa interpretacion, y además el insultado resulta que eres tu!
Ya te lo dije una vez ; Dejanos Tranquilos!
Yo por mi parte cierro mi participacion en este foro hasta que se normalice.
Saludos
Etpour... y sin embargo se muve... ya lo dijo Galileo camino de su destierro a quienes no quisieron ver la realidad.
Hay 6 comentarios.
1