Autor: aladelta
viernes, 21 de abril de 2006
Sección: Antropología
Información publicada por: aladelta
Mostrado 268.977 veces.
La genética y su aplicación en el estudio de las poblaciones humanas.
Se da un repaso sobre las últimas teorías sobre el origen del hombre moderno, para luego ahondar en los estudios de genética de poblaciones, haciendo hincapié en el uso del ADN mitocondrial y el cromosoma-y. Se centra finalmente y sobre todo en la población europea y muy especialmente en las poblaciones ibéricas.
Mayormente son copias de otros documentos o traducciones con algún pequeño comentario mío. Creo que afortunadamente hay en Celtiberia.net un creciente interés sobre el estudio de poblaciones mediante la genética, y espero que este articulo de una idea mejor y ayudar a interpretar mejor los documentos publicados. Ojalá se anime más la gente para que este no sea el único articulo sobre este tema.
Si alguno de vosotros desea pasarme información sobre el tema o incluso corregir algo , enviadme un correo a aladelta@yahoo.com
Saludos.
- LAS TEORÍAS SOBRE EL ORIGEN DEL HOMBRE MODERNO.
- ---EVIDENCIAS APORTADAS POR GEÓLOGOS: LA SUPEREXPLOSIÓN DEL TOBA.
- ---CONCLUSIÓN:
- ---¿SOMOS DESCENDIENTES DEL NEANDERTAL?.
- ---EL HOMO FLORESIENSIS.
- ALGUNOS CONCEPTOS QUE APARECEN COMÚNMENTE EN LAS PUBLICACIONES DE ESTUDIOS GENÉTICOS DE POBLACIONES.
- LAS HERRAMIENTAS REY DEL ESTUDIO DE LAS POBLACIONES: EL ADN MITOCONDRIAL Y EL CROMOSOMA-Y.
- ---EL ADN MITOCONDRIAL.
- ---------EL HOMBRE DE CHEDDAR.
- ---------OETZI, EL HOMBRE DE HIELO.
- ---------LAS SIETE HIJAS DE EVA.
- ---EL CROMOSOMA-Y.
- ---------VARIACIÓN EN EUROPA DE HAPLOGRUPOS DE CROMOSOMA-Y.
- EL MODELO DE DIFUSIÓN DÉMICA.
- COMPOSICIÓN GENÉTICA DE LA PENÍNSULA IBÉRICA.
- ---COMPOSICIÓN DESDE LA PERSPECTIVA DEL ADN MITOCONDRIAL.
- ---COMPOSICIÓN DESDE LA PERSPECTIVA DEL CROMOSOMA-Y.
- FENOTIPOS, LENGUAS Y GENES.
- ---FENOTIPOS.
- ---LENGUAS.
- BIBLIOGRAFÍA.
>LAS TEORÍAS SOBRE EL ORIGEN DEL HOMBRE MODERNO.
1)La Teoría Multirregional
(Wolpoff et al. 1984)
2)La Teoría del orígen africano
(Cann et al. 1987, Stringer y Andrews 1988)

El modelo multirregional propone que el hombre moderno tiene su origen en diferentes poblaciones a partir del homo erectus, oriundo de África pero con una antiguedad de hace 2 millones de años. Estas poblaciones diferenciadas a partir de tan alejado momento daría lugar, por separado, al hombre moderno, coincidiendo con las diferentes razas actuales.
La Teoría del orígen africano es en realidad la del origen africano RECIENTE, o la Teoría de Fuera de África, la cual propone que un descendiente del homo erectus, el hombre moderno surgió de África hace unos 150.000 años y pobló todo el planeta, sustituyendo las diferentes poblaciones también descendientes del homo erectus, pero sin señales de hibridación. Una versión modificada de esta teoría es la que propone que después de la salida de África se produjo una reducción drástica de la población debido a algún tipo de evento extraordinario y que se expandió a partir de ese cuello de botella hace 50.000 años de nuevo desde África(segunda emigración). Es la llamada weak garden of eden hypothesis (WGE). Los genetistas demógrafos creen que la población humana ancestral era muy reducida --unas escasas 2.000 personas en condiciones de reproducirse, según estimaciones publicadas en diciembre de 1999.

EVIDENCIAS APORTADAS POR GEÓLOGOS: LA SUPEREXPLOSIÓN DEL TOBA.
El Toba fue un supervolcán que se encontraba en Sumatra y hace 74.000 años su caldera explotó de la forma más violenta que ha existido. Su explosión está calificada con las siglas VEI8, el punto máximo de la escala de erupciones volcánicas. Esta caldera tenía aproximadamente la superficie que tiene ahora el lago que dejó en su lugar, unos 100 km de largo y 60 km ancho. El Toba emitió gran cantidad de partículas a la atmósfera. Esto supuso un cambio climático en la Tierra durante muchos años. La radiación solar no podía llegar a la superficie porque la estratosfera estaba totalmente colapsada de particulas. La luz no penetraba para alimentar a las plantas y los animales (entre ellos nosotros) no podían alimentarse de ellas ni de otros animales porque escaseaban. Los veranos se hicieron más frescos, la nieve no se derretía y se acumulaba para el invierno siguiente, y así sucesivamente.
 |  |
|---|

El geólogo Michael Rampino estimó, con los datos acumulados por explosiones conocidas y registradas en la actualidad, que tras la superexplosión del Toba la temperatura global del planeta descendió 5ºC. Eso supuso para Europa que sus veranos sufrieran un descenso de 15 ºC. Lo cierto es que los genetistas andaban como locos intentando averiguar qué le pasó a nuestra especie para que ésta casi desapareciera hace entre 80.000 y 70.000 años. Aquí encontramos la razón. Fue un antropólogo, Stanley Ambrose, quien relacionó la explosión del Toba con la casi extinción de nuestra especie. Por supuesto que cuando ocurrió este evento, las poblaciones ya habían comenzado a colonizar el planeta y de echo se cree que los Aboríenes australianos ya habían iniciado su camino hacia su actual ubicación.
CONCLUSIÓN.
La Teoría multirregional fue propuesta por paleontólogos y está sustetada por el registro fósil, mientras que a la Teoría del origen africano reciente la respalda la evidencia de los estudios genéticos cuyos resultados le da un amplio margen a su favor. A demás, las Teorías basadas en el registro fósil tiene la problemática de ser incompletas, llenas de huecos difíciles de interpretar. Por otra parte es difícil descartar completamente una Teoría multirregional más compleja con intercambio entre poblaciones, aunque para que esto pudiera haber sido así, tendría que haber habido un flujo de genes suficientes entre poblaciones geográficamente muy distanciadas a lo largo de 2 millones de años, y para ello la población debería de haber sido muy grande, permitiendo ese flujo genético entre las poblaciones, lo cual la evidencia del registro fósil no apunta en esa dirección. De todas formas, como veis ambas teorías proponen el origen del hombre moderno en África, aunque difieren en el momento en que esta emigración se llevó a cAbo.
¿SOMOS DESCENDIENTES DEL NEANDERTAL?.

Los neandertales evolucionaron durante cientos de miles de años en Europa, y se apunta a que quedaron aislados del resto de homínidos, mientras que nuestros antepasados se cree que todavía no habían salido de África.

Sobre si el neandertal y nuestra especie pertenecían a la misma especie se realizó un estudio en el cual se pudo extraer una muestra de ADN de un neandertal apta para el análisis y compararlo con los del hombre moderno. El ADN no fosiliza y, aunque lo encontremos, en la mayoría de las veces estará contaminado. Aún así, se pudo extraer una muestra de ADN "sin contaminar" del interior de un hueso fosilizado de neandertal y analizarlo. El resultado de este análisis demostró que su ADN es muy parecido al nuestro, pero distinto, demostrando que somos dos especies emparentadas(ambas especies descienden del homo antecessor) pero diferenciadas por el aislamiento mútuo. Nuestro linaje y el de los neandertales se separó hace entre 550.000 y 690.000 años, mientras que nuestra diversidad actual surge entre hace 120.000 y 150.000 años.


Un híbrido potencial Neandertal/sapiens de hace 24,500 años proveniente del sitio Lagar Velho, en Portugal, fue anunciado.
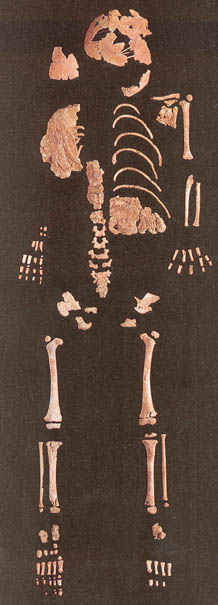
Este individuo de 4 años de edad tiene un cuerpo corto y ancho como un Neandertal, pero posee un cráneo anatómicamente moderno. Existe una serie de problemas en la interpretación de este hallazgo como un híbrido Neandertal/sapiens. Primero que todo, como un híbrido, debería tener una mezcla de caracteres en todo el cuerpo y no simplemente poseer el cuerpo de un Neandertal y el cráneo de un humano moderno. Por ejemplo, si observamos a los híbridos entre leones y tigres, ellos no poseen la cabeza de una especie y el cuerpo de la otra, sino que exhiben una mezcla morfológica de ambas especies. Segundo, y más importante, la aceptación de este espécimen como un híbrido sugeriría que los caracteres de los Neandertales fueron retenidos por entre 6,000 a 10,000 años después de que los Neandertales se extinguieron, lo cual es muy poco probable. Esto es teóricamente improbable porque los caracteres de los Neandertales habrían sido completamente diluidos por los genes de Homo sapiens en un periodo de tiempo tan largo.
La paleontología y la genética, de todas formas, parece dejar la puerta abierta a ésta hibridación entre neandertales y hombres modernos, pero para demostrar eso se deben encontrar fósiles y conseguir ADN no contaminado para compararlo con la de los hombres modernos, y de momento no se ha conseguido.
Enlace a EL ÚLTIMO NENDERTAL
Para saber más sobre el niño de Lagar Velho (Portugal) pincha aquí
EL HOMO FLORESIENSIS.

En septiembre de 2003 científicos indonesios y australianos hicieron un increíble descubrimiento en la isla de Flores, Indonesia. Encontraron enterrados en una cueva un esqueleto de lo que parecía un niño, por su poca estatura, alrededor de un metro. Sin embargo, después de examinarlo, se dieron cuenta de lo que tenían delante no era sino un homínido adulto hembra. Los restos tienen una edad de 18.000 años y tras examinarlos detenidamente se llegó a la conclusión de que era una especie nueva descendente probablemente de una variante del homo erectus que se estima llegó a la isla hace aproximadamente un millón de años evolucionando en ella hasta alcanzar estas características. Era bípedo y caminaba erguido, usó herramientas de piedra sofisticadas, y parece que hasta supo hacer fuego, el cual utilizaba para cocinar la caza de la que se alimentaba, ¡todo eso con una capacidad craneana de 380 cc, parecido al tamaño del cerebro de chimpancé!, lo cual desmonta la teoría de la necesidad de un cerebro grande para el desarrollo de la inteligencia. Se cree que se extinguió hace unos 12.000 años a raíz de una erupción volcánica que devastó la isla, o que perdió la batalla contra otros grupos homo.

Lo más intrigante de todo esto es que los habitantes de la isla cuentan leyendas de unos seres de las mismas características que llamaban Ebu Gogo, los cuales se murmuraban los unos a los otros en lo que parecía un lenguaje, repitiendo como loros lo que decía el otro, a modo de colación. ¡La última leyenda data de 100 años!. Henry Gee, el editor de la revista Nature se atreve incluso a especular que estas criaturas puedan todavía existir en algún lugar de zonas vírgenes de indonesia.

Alguien podría pensar si el homo floresiensis tiene algo que ver con los pigmeos. Rotundamente no. Los pigmeos pertenecen a nuestra misma especie, son nuestros parientes directos, nuestros hermanos de sangre; son señores bajitos y cabezones, con una capacidad craneana comparable a la del resto de humanos modernos. Mientras que el homo floresiensis es nuestro primo lejano, que aunque vivió al mismo tiempo que nuestra especie pertenece a una especie inteligente, pero diferenciada de la nuestra.

El principio de coalescencia: Asume la existencia de un origen común a todos los seres vivos. Por este mismo principio se asume un origen común a todos los seres humanos.
Marcadores clásicos: Las investigaciones se centraban en el estudio de antígenos eritrocitarios (sistema Abo, Rh, MN), proteínas séricas, enzimas eritrocitarias y sistema HLA.
RFPL (Restriction Fragment Length Polimorphism):Análisis de polimorfismos de longitud de fragmentos de restricción.
HLA (human leukocyte antigen): Antígenos de los leucocitos humanos. Determinan la compatibilidad de los transplantes.
LGM (Last Glacial Maximum): La máxima expansión de los hielos durante la última glaciación se calcula que ocurrió en el 18.000 a. C. aproximadamente. La última glaciación o glaciación de Würm ocurrió entre el año 35.000 a. C. Y el 10.000 a.C aprox.
Microsatélites o STRs (Short Tandem Repeat polymorphisms). Son secuencias de ADN repetidas en tandem de 2 a 6 pares de bases. Se emplean como marcadores genéticos para rastrear la herencia familiar o mapear enfermedades en el genoma. La tasa de mutación es grande, por lo que se utiliza para conocer eventos demográficos recientes ocurridos en una escala de tiempo más reducida.
Minisatélites o VNTR (variable number of tandem repeats). Son repeticiones también en tandem. Se diferencian de los microsatélites en que las secuencias que se repiten son mayores.
Polimorfismos de inserción alu. Las inserciones Alu son elementos polimorficos de aproximadamente 300 pares de bases, que se encuentran distribuidas en el genoma de los primates. La especie humana posee incorporada en sus celulas su propia subfamilia. La inserción polimórfica en lugares específicos del genoma de estos elementos Alu, sirven para el estudio de la historia demográfica de las poblaciones. Sin embargo el grado de polimorfismo es bajo (sólo hay dos alelos, que en realidad son presencia y ausencia de inserciones en las secuencias). Tampoco se pueden construir filogénias, por lo que se limita su aplicación para el estudio de poblaciones.
Mutaciones puntuales o SNPs (Single Nucleotid Polymorphism). Un SNP se define como la presencia de dos posibles bases en una posición particular en el ADN. Esta posición particular debida a mutaciones puntuales constituyen la principal contribución a la variación genética humana y cuya densidad en el genoma es del orden de 0,5-10 por 1000 pares de bases. El interés de los SNPs reside en su posible asociación con el desarrollo de enfermedades y en las enormes posibilidades que ofrecen para el descubrimiento de la base genética de la susceptibilidad a las enfermedades complejas o de la sensibilidad a los agentes terapéuticos (farmacogenética). Marcador bialélico. Asociado con la porción del cromosoma-y que no se recombina en la fecundación.
Alelos de diferentes genes (HLA entre otros). Un alelo es una de las formas variantes de un gen en un locus(lugar del cromosoma donde está localizado un gen específico) o de un marcador particular en un cromosoma. Diferentes alelos de un gen producen variaciones en las características hereditarias tales como el color del cabello o el tipo de sangre.
NRY: Non-Recombining region of Y chromosome. Región del cromosoma-y que no se recombina durante la fecundación.
Fenotipo: El fenotipo de los organismos comprende las características observables directamente por nuestros sentidos(morfología), que se originan como consecuencia de las interacciones entre el genotipo y el ambiente.
Alelo dominante y alelo recesivo: Son dos formas de un mismo gen que codifican para caracteres determinados. El alelo dominante es el que, estando en uno solo de los cromosomas de cada pareja de un individuo, se manifiesta en el fenotipo de éste. El recesivo es el alelo que debe estar presente en los dos cromosomas para que se manifieste en el fenotipo el carácter para el que codifica.
ADN (el): Abreviatura de ácido desoxirribonucleico, constituyente esencial de los cromosomas del núcleo celular.
Genoma: El material genético de un organismo.
Mitocondria: En el citoplasma de las células con núcleo diferenciado, orgánulo encargado de la obtención de energía mediante la respiración celular.
Cromosoma: Cada uno de los filamentos de material hereditario que forman parte del núcleo celular y que tienen como función conservar, transmitir y expresar la información genética que contienen: El ser humano tiene 23 pares de cromosomas.
Haploide(adj): Referido a un organismo o a su fase de desarrollo, que tiene una dotación simple de cromosomas. Los óvulos y los espermatozoides son células haploides, pero un embrión es ya diploide.
Filogenia: La evolución de un grupo de organismos genéticamente relacionados, para diferenciarla del desarrollo de un organismo individual.
Marcador: También conocido como marcador genético, es un segmento de ADN cuya herencia se puede rastrear. Un marcador puede ser un gen, o puede ser un segmento de ADN sin función conocida. Dado que los segmentos de ADN que se encuentran contiguos en un cromosoma tienden a heredarse juntos, los marcadores se usan a menudo como formas indirectas de rastrear el patrón hereditario de genes que no han sido aún identificados, pero cuyas ubicaciones aproximadas se conocen.
Haplotipo: tipo de secuencia que comprende todas las secuencias idénticas. Estas secuencias están compuestas por combinaciones alélicas.
Haplogrupo:grupo de haplotipos que comparten un ancestro común. Es decir, estos haplotipos forman un conjunto que se repite en una población, y que define el haplogrupo.
Hg: Haplogrupo
MtDNA: Ácido desoxiribonucléico(ADN) mitocondrial o ADNmt.
Me dejo otras muchas definiciones, pero “grosso modo” están las más importantes para tener una idea general.
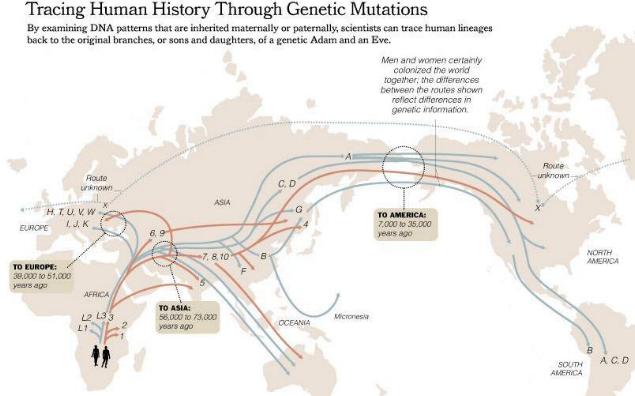
Los linajes que resultan como consecuencia de mutaciones, tanto en el ADN mitocondrial como en el cromosoma-y son conocidos como haplogrupos. Cada haplogrupo lo conforman diversos haplotipos que lo definen.
Las mutaciones que producen la variación de haplotipos dan como resultado un nuevo subhaplogrupo. Todos los seres humanos compartimos haplotipos comunes, que definen el haplogrupo común, nuestro Adam del cromosoma-y, o nuestra Eva del ADN mitocondrial. A partir de éste haplogrupo común surgen otros haplogrupos diferenciados por variaciones de estos haplotipos, y así sucesivamente.
EL ADN MITOCONDRIAL.

El ADN mitocondrial no se encuentra en el núcleo de las células, sino en unos orgánulos del citoplasma (es decir, fuera del núcleo) que se llaman mitocondrias. Cuando se produce la fecundación del óvulo, el espermatozoide aporta la mitad de los cromosomas. La otra mitad los pone el óvulo. Pero sólo la madre proporciona el ADN mitocondrial, ya que el espermatozoide no contribuye con mitocondrias(la cola del espermatozoide posee mitocondrias, pero se pierde durante la fecundación del óvulo).
En principio, todas las personas deberían tener la misma cadena de letras de ADN en sus mitocondrias. En la realidad, el ADN mitocondrial ha acumulado progresivamente cambios durante milenios debido a errores de copia y a daños por radiación. Es por eso que algunos cambios sólo aparecen en regiones o continentes específicos. En un artículo publicado en marzo del 2000 en The American Journal of Human Genetics, Douglas C. Wallace y algunos colegas identifican a los Vasikela Kung, del noroeste del desierto de Kalahari, al sur de África, como el grupo racial que está más próximo a la raíz del árbol del ADN mitocondrial humano. Otro grupo racial que parece casi igualmente antiguo es el de los pigmeos Biaka del África Central.
El árbol del ADN mitocondrial está enraizado en un único individuo, la Eva mitocondrial, porque todos los otros linajes se extinguieron.
El primero en utilizar la Herramienta del ADN mitocondrial fue Wesley Brown en 1980. Pero el que realmente tuvo repercusión popular fue el equipo de Allan Wilson en 1987. Ambos, sin embargo, llegaron a las mismas conclusiones, esto es: el origen común de todos los seres humanos en un linaje no más antiguo que 180.000 años. Luego vendrían otros complementando a los anteriores, el grupo de Cann en el 87 con su Eva mitocondrial basado en el principio de coalescencia, Vigilant y colAboradores en el 91, corroborando el anterior, así hasta más tarde desentrañar los diferentes haplogrupos a partir del original.
Dejando de nombrar ya un poco a los muchos autores que han estudiado y definido los diferentes haplogrupos, empezaremos a poner nombre a estos:
Macrohaplogrupo L

Pinchar aquí para ver la expansión de mtDNA mundial según J. D. Mcdonald, por pasos.
(nuestra Eva mitocondrial común), que ha su vez se subdivide en L0, L1, L2, L3, M y N. Los subhaplogrupos L0, L1, L2 son específicos del África subsahariana, mientras que los M y N aparecidos al noreste de África, se expandieron por Europa y Asia.
Los haplogrupos que poblarán Asia surgen tanto del macrohaplogrupo M (que son los denominados C, D, E, G, Z) y el macrohaplogrupo N (subdividido en A, B, F, Y)
Los haplogrupos que poblarán América (antes de las invasiones europeas) son el A, B, C, D y X constituyendo el 100% de linajes.
Los haplogrupos que poblarán Europa surgen del macrohaplogrupo N y son: H, I, N1b, T, U, V, W y X. Estos haplogrupos constituyen el 98% de los linajes europeos.
EL HOMBRE DE CHEDDAR.

En 1903, en una cueva cerca de Cheddar, en el sur de Inglaterra, se encontró enterrado un esqueleto no fosilizado de un hombre. Se determinó que el esqueleto tenía una antigüedad de 9.000 años. Bryan Sykes analizó una prueba de ADN mitocondrial de una muela del esqueleto. Su ADN fue comparado con el de los habitantes de Cheddar. Los resultados fueron impactantes. El hombre de Cheddar, que había vivido hacía 9.000 años, resultó tener un pariente directo vivo en Cheddar, un profesor de escuela. El test también sirvió para demostrar que los britones no eran descendientes de emigrantes neolíticos venidos de Oriente Medio, como se había creído tradicionalmente, sino que estos eran descendientes de cazadores recolectores que llegaron a las islas hace aproximadamente 10.000 años.
OETZI, EL HOMBRE DE HIELO.
En 1991 dos turistas alemanes encontraron lo que parecía los restos de un ser humano momificado y conservado por deshidratación, el frío y la nieve que lo cubría, que se tornó en hielo durante los más de 5.000 años que estuvo enterrado en él; estaba atrapado en hielo glaciar, en los Alpes tiroleses, cerca de la frontera entre Alemania y Austria. Junto al cuerpo había un hacha de cobre, una daga, un arco y 14 flechas. El estudio forense evidenció que el individuo al que se estimó de 46 años de edad en el momento de su muerte y de 1.59 m de altura, fue herido de muerte por una flecha que le entró a la altura del pecho y murió poco después desangrado. Algunos “alumnos aventajados” dijeron que se trataban de un fraude, una momia india americana falsa, plantada a propósito en el hielo glaciar. Así que tras remover tejido del cuerpo por los forenses para ser analizado mediante la prueba del ADN mitocondrial por el equipo del profesor Bryan Sykes se llegó a la conclusión de que no estaba emparentado ni con ningún nativo americano, ni con siberianos, ni subsaharianos, sino que lo estaba con los habitantes que habitaban hoy en día en los Alpes. Un grupo de investigadores de la Universidad de Queensland, Australia, determinó que el hombre de hielo nació en lo que ahora es el pueblo de Velturno(Feldthurns), en el sur del tirol Italiano (norte de Italia). Se ha encontrado incluso un pariente actual de Oetzi, que vive en Dorset, sur de Inglaterra; es una irlandesa que dio la casualidad que trabajaba en el lAboratorio de Sykes. Los científicos, además, lograron precisar que el hombre de hielo pasaba los veranos en las montañas de Vinschgau, al norte de su pueblo natal.
 |  |
|---|
La conclusión a la que se llegó, gracias a la prueba genética y la del carbono 14, (isótopos hallados en los huesos y dientes) y la composición química de muestras de agua y tierra de un área de los Alpes, fue que el hombre de hielo, al que llamaron Oetzi, era un cazador neolítico que murió intentando cruzar los Alpes hace 5.300 años y que esta emparentado con los habitantes actuales de esa zona, además de con el resto de europeos, lo que además demuestra la estabilidad del ADN para su aplicación al estudio de poblaciones.
Para saber más, pincha aquí.
LAS SIETE HIJAS DE EVA.

A raíz del estudio de Oetzi, Sykes decidió, movido por la gran curiosidad de encontrar parentescos entre los seres humanos, dejar su carrera de genetista médico para concentrarse en el estudio de las poblaciones.
En el 2001 publicó “las siete hijas de Eva”. En él, además de hablarnos del hombre de hielo, nos revela que a partir de sus estudios realizados durante una década, ha descubierto que la mayoría de europeos desciende de sólo siete linajes de ADN mitocondrial, o como a él le gusta llamarlas, las siete hijas de Eva. Además les puso nombres inspirados en la mitología europea:
Ursula(osa en latín): Se corresponde con el haplogrupo U5. Es el linaje más viejo de los siete. Se remonta a 45.000 años con los primeros humanos modernos. El 11% de los europeos descienden directamente de ella. Extendido por toda Europa, es especialmente frecuente en el oeste de las Islas Británicas y Escandinavia.
Xenia(hospitalario en griego): Se corresponde con el haplogrupo X. Es el segundo linaje más antiguo de los siete. Fue fundado hace 25.000 años por la segunda oleada de hombres modernos, justo antes de la llegada de la última glaciación. El 7% de los europeos pertenecen a este linaje. Se subdivide en tres subhaplogrupos; uno extendido por el Este de Europa y los otros dos por el Centro. Curiosamente el 1% de los indígenas americanos forman parte de este grupo.
Helena(luz en griego). Se corresponde con el haplogrupo H. Es, con diferencia, el linaje más extendido en toda Europa con un 41% de europeos que pertenecen a este linaje. Este se originó hace 20.000 años en algún lugar entre los valles de Dordoña y el norte de la Península Ibérica. Como ya he dicho, está extendido por toda Europa, aunque su frecuencia más alta está entre los vascos. Su expansión se inició después de la retirada de los hielos, al finalizar la última glaciación.
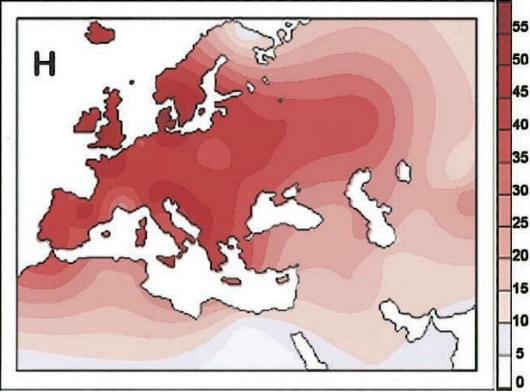
Velda(soberana en escandinavo). Se corresponde con el haplogrupo V. Es el linaje más modesto de los siete con sólo un 4% entre europeos descendientes directos. Velda vivió inicialmente en el norte de la Península Ibérica hace 17.000 años. Sus descendientes habitan principalmente en el oeste y norte de Europa siendo curiosamente muy frecuentes entre los lapones, habitantes de Finlandia y norte de Noruega.
Tara(colina rocosa en gaélico). Se corresponde con el haplogrupo T. Aproximadamente el 10% de los europeos pertenece a este linaje. Se expandió desde el Norte de Italia entre las colinas toscanas y las rías del río Arno hace 17.000 años. Su principal expansión se centra en el sur y oeste europeos, alcanzando grandes concentraciones en Irlanda y oeste de Gran Bretaña.
Katrine(pura en griego). Se corresponde con el haplogrupo K. El 10% de los europeos pertenece a este linaje. Vivió hace 15.000 años en las planicies boscosas del Noreste de Italia, hoy en día inundada por el Adriático, y entre las estribaciones de los Alpes. Sus descendientes residen mayoritariamente en ese mismo sitio, pero también se han extendido bastante hacia el centro y norte de Europa.
Jasmine(flor en persa). Se corresponde con el haplogrupo J. Es el segundo linaje más extendido en entre la población europea, después de la prolífica Helena. También es el único linaje que tiene su origen fuera de Europa. El 12% de europeos pertenece a este linaje. Tiene su origen en el Oriente Medio hace 8.500 años. Los pertenecientes a este linaje son los descendientes de los primeros agricultores que expandieron la agricultura desde Oriente Medio.
Bryan Sykes concluye afirmando que el 80% de los Europeos descienden de sus antepasados paleolíticos.
Después se ha ido añadiendo más “hijas europeas de Eva”, menos importantes por su poca propagación por Europa. Las nuevas son:
Ulrike(Señora de todo en alemán). Se corresponde con el haplogrupo U4. Con menos de un 2% de descendientes. Originaria del frío refugio que fue Ucrania, hace 18.000 años. Sus descendientes se encuentran mayoritariamente en el este y norte de Europa, con altas concentraciones en Escandinavia y los países bálticos.
Iris(del griego, con nuestro mismo significado de arco iris). Se corresponde con el haplogrupo I. Posee menos del 2% de descendientes. Se originó aproximadamente hace 43.000 años en algún lugar de Irán, siendo los Kurdos los máximos representantes. Se propagó por Europa durante la venida de emigrantes portadores de la cultura gravetiense, hace unos 25.000 años.
JT. Hoy en día se cree que T (Tara)y J(Jasmine) vinieron “juntas” de Oriente Medio y son los principales protagonistas de la expansión de la agricultura en Europa. También va cogiendo más fuerza que estos grupos trajeron las lenguas indoeuropeas a la Europa sur y central, primero, y más tarde a la occidental y norte.
Bryan Sykes se fijó especialmente en el pueblo vasco, tradicionalmente considerado como el más antiguo de Europa. Su análisis genético no demostró nada peculiar: ofrecían los mismos resultados que los de todos los europeos, pero con una excepción notable: había representantes de los seis grupos más antiguos, pero no encontraron ninguno del más moderno, el correspondiente a Jasmine. Además son el grupo que más individuos pertenecen al linaje de Helena.
EL CROMOSOMA-Y.
ver expansión 1

ver expansión 2
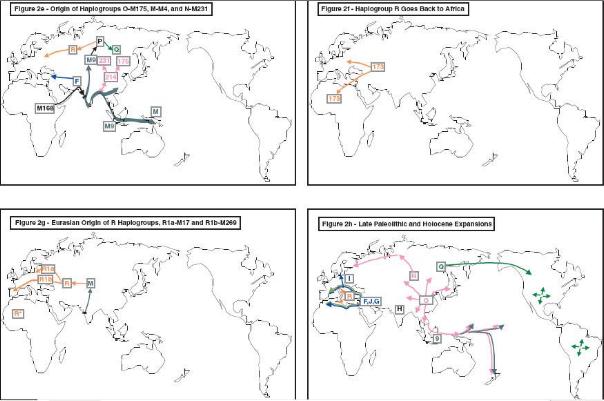
El cromosoma-Y tuvo un protagonismo más reciente. Durante mucho tiempo fue considerado como un cromosoma no muy polimorfico, consistente en grandes cantidades de lo que se conoce como ADN basura, conteniendo muy pocos genes. Sólo durante esta última década muchos nuevos estudios del cromosoma-y ha mejorado sustancialmente el conocimiento sobre diferentes aspectos en lo que se refiere a su estructura y función. Su específica característica importante para el estudio de poblaciones, por la presencia de grandes áreas que no se recombinan durante la fecundación, manteniendo una transmisión únicamente patrilineal, sirve como instrumento para muchos campos de la biomedicina. El cromosoma-y también ha acumulado progresivamente cambios durante milenios debido a errores de copia y a daños por radiación.
El árbol del cromosoma-y está igualmente enraizado en un único individuo, el Adam del cromosoma-y.
ver arbol


La razón por la que el cromosoma- y es una herramienta apropiada para investigar la evolución humana reciente, genética médica y reconstruciones genealógicas, es por la singularidad que presenta entre los otros cromosomas humanos. Tiene un papel determinante en la herencia sexual, pasando únicamente de padre a hijo varón.
Los primeros estudios en este campo se iniciaron en 1985 cuando fue desvelado el primer polimorfismo en el cromosoma-y (Casanova et al. 1985).
En los 90 fueron descubiertos nuevos polimorfismos (“Fathers and sons: the Y chromosome and human evolution” Jobling, M.A. and Tyler-Smith, C. (1995)). Pero Sólo durante esta última década, arrancando con la publicación en el 1997 de Peter A. Underhill y en particular durante los últimos 5 años, ha sido cuando este campo ha tenido gran desarrollo.
Después de su comprobada gran utilidad para el estudio de poblaciones, el estudio del cromosoma-y ha ido incrementando y generalizando por grupos de estudio. Así, muchos grupos empezaron a estudiar poblaciones mediante esta herramienta y se han llegado a utilizar al menos siete diferentes nomenclaturas. Todas ellas tienen sus símbolos propios y son equivalentes. Al ser complicado seguir tan variada nomenclatura el consorcio del cromosoma-y (YCC) ha desarrollado una nomenclatura única para posteriores publicaciones.
Las dos ramas más viejas son A y B. Ambas muestran una gran distribución en el África subsahariana, aunque presentan moderadas o bajas frecuencias. La razón es la expansión que supuso sobre todo el continente africano del haplogrupo E. El 80% de africanos es descendiente directo de este linaje.
A y B están restringidos a África.
La mutación M168 representa la firma de la moderna emigración desde África al resto de continentes y África misma corroborando la teoría del origen africano reciente, excepto para los haplogrupos Ay B, restringidos en África, ya en bajas proporciones en ese continente. M168 representa, pues, el verdadero árbol coalescente.
La mayoría de ramas del árbol que representan los haplogrupos del cromosoma-y son los Hg C, D, E, y F.
La mutación que generó C parece haberse producido fuera de África, pues sus descendientes sólo habitan fuera de esta. Es probable que se generara en Asia, expandiéndose a América desde el centro y norte y a Oceanía desde el sureste.
Los haplogrupos E y D comparten su origen en África. Algunos descendientes de estos linajes permanecieron en el continente primigenio, dando lugar al haplogrupo E, el mas frecuente en africa como ya se ha dicho antes.
El Haplogrupo D se extiende por Asia aunque con bajas frecuencias, con la excepción de zonas periféricas como el Tibet, Japon e Islas Andaman.
El superhaplogrupo F, caracterizado por la mutación M89, es el padre del resto de los siguientes haplogrupos:
J y G en Oriente Medio, I en Europa, H en el sudeste de Asia. La mutación M9 da como resultado otro nuevo gran linaje llamado K. Las ramas de este gran subgrupo migraron en varias direcciones, (norte y este, principalmente) dando lugar a nuevos subhaplogrupos a partir de este haplogrupo K.
Hg J probablemente tiene su origen en Oriente Medio y es ahí donde alcanza su máxima frecuencia y diversidad, disminuyendo hacia el mediterráneo europeo, norte de África, Irán, Asia central y la India. Se divide en dos subhaplogrupos: J1 y J2, los cuales son los más comunes de nuevo en Oriente Medio. La distribución de los cromosomas-y sugiere que el hg J2 se originó en la parte norte, asociado a la expansión desde Anatolia a la parte sudeste de Europa, reflejando una expansión de agricultores desde Anatolia; y el hg J1 en la parte sur, asociado con la difusión semita(árabes, fenicios, sirios...a excepción de judios y palestinos, más parecidos a la parte anatolia, con más porcentaje de J2.)
Así del K surgen: L con grandes frecuencias en el sudeste de Asia. M restringido en Australia y Nueva Guinea. O Predominando en zonas del sur y sudeste de Asia, alcanzando el norte de China, Manchuria y algunas poblaciones siberianas. P que genera Q y R en Eurasia; Q, es característico en la población siberiana y amerindia; R es característico de la población europea y el oeste de Asia.
Por supuesto, después de esta primera expansión, cada región continental ha desarrollado su propia rama especifica; por ejemplo del linaje R resultaría R1, y R2; de R1 resulta en R1a y R1b, etc.
Además el oeste de Asia y Europa han recibido una ola adicional de genes procedentes de Africa, presumiblemente vía corredor levantino (Oriente Medio), trayendo linajes E, ausentes en la India.
VARIACIÓN EN EUROPA DE HAPLOGRUPOS DE CROMOSOMA-Y.
ver mapa
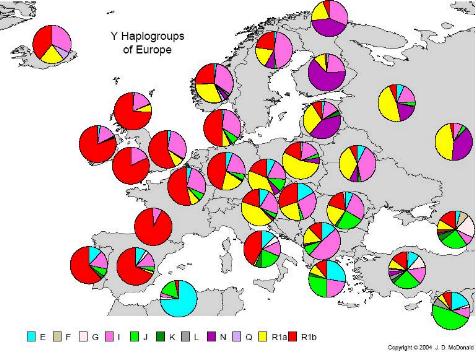
La población europea ha sido una de las que más estudios a disfrutado en los últimos años. En el año 2.000, Ornella Semino y colAboradores publicó el mayor estudio que se había efectuado hasta el momento en la población europea. Aunque Rosser y colAboradores hicieron un estudio paralelo el mismo año consiguiendo resultados similares, el estudio de Semino tuvo más eco en los medios. En el estudio de Semino resultó que más del 95% de los cromosomas-y estudiados en europeos pueden ser agrupados en 10 haplogrupos. La distribución geográfica y estimaciones de tiempo desvelaron que la mayoría de europeos desciende de dos oleadas paleolíticas y una neolítica.
La mayoría de europeos pertenecen a los Hgs R1a, R1b, I y N3, (grupos paleolíticos los cuales cubren el 73% del total) y a los Hgs J2, E3b y G (grupo neolítico que cubren el 27% restante).
La distribución de los linajes a partir de la mutación M173, sugiere que es un antiguo marcador eurasiático traído por el primer grupo de homo sapiens sapiens que entró en Europa y difundió la cultura auriñaciense hace entre 40.000 y 35.000 años. La estimación de estos linajes ha sido corroborada en estudios posteriores, tanto genéticos como arqueológicos. El 50% de europeos comparten el marcador M173 que define hg R1 y que dio lugar a los subhaplogrupos R1a, y R1b(o R1*).El hg R1b presenta altas frecuencias en el oeste europeo, siendo la Península ibérica, Irlanda, y Reino unido los máximos representantes, decreciendo hacia el este, mientras que el hg R1a presenta una frecuencia opuesta al anterior con una frecuencia máxima en el este europeo, especialmente en la población eslava.
Se atribuyó la diseminación del grupo R1a a la recolonización postglaciar, iniciada hace unos 11.000 años, desde el refugio glaciar ucraniano, aunque recientemente se ha querido atribuir la expansión por la emigración Kurgan desde el mar Caspio, siendo esta expansión entonces más reciente, según Rosser y colAboradores.
El grupo R1b cubre un área mayor que Europa, pero, como ya he dicho, el origen de este está en el occidente Europeo, revelando una diseminación postglaciar desde Iberia, lugar que sirvió de refugio para este linaje.
La mutación M170, que da lugar a hg I, revela otra expansión paleolítica, estimada de hace unos 22.000 años. Son los descendientes de hombres que llegaron desde Oriente Medio hace unos 25.000 años, asociados con la llegada de la cultura gravetiense. Los Balcanes fueron el refugio para la mayoría de este grupo.
N3 Está presente en el norte y este europeos, pero ausentes en el oeste y sur. Es también frecuente en el norte de Asia.
Los marcadores M35, M172, y M201, dan como resultado los haplogrupos E3b, J2, y G respectivamente y su frecuencia decrece desde Oriente Medio a Europa, por lo tanto estos han sido considerados los representantes de la contribución masculina de la difusión démica de agricultores venidos de Oriente Medio. Además ese flujo genético es más pronunciado en las costas mediterráneas que en la Europa continental.
En 1984 Ammerman AJ. Y Cavalli-Sforza LL. Publicaron “The Neolithic Transition and the Genetics
of Populations in Europe”, la culminación de un trabajo de más de diez años. En ella exponía el modelo de difusión neolítica. Para ello se basó en el estudio genético basado en Frecuencias de alelos en el que detectó un cambio gradual de las frecuencias de genes de este a oeste. A partir de ahí dedujo que se había producido un flujo de genes desde el Próximo Oriente en todas direcciones en época neolítica, el cual resultó coincidir con los datos arqueológicos del origen de la agricultura, llevando a pensar a Ammerman y Cavalli-Sforza que la estructura fundamental de la población europea fue determinada por la dispersión neolítica, e idearon el modelo que llamaron de difusión démica, el cual se define como el proceso específico de expansión geográfica de un grupo desde un lugar origen desencadenado por una gran innovación o innovaciones(agricultura, organización, avances de tipo militar...) que permiten o estimulan el crecimiento de una población y la expansión hacia nuevas áreas. Esta difusión se realizaría siguiendo el modelo denominado OLA EN AVANCE, el cual establece que la agricultura fue transmitida por movimientos de campesinos, que al aumentar en población a un ritmo 50 veces superior al de los cazadores-recolectores, irían extendiéndose buscando nuevas tierras para sus cultivos.

En realidad faltaba por saber la magnitud de esa expansión de genes, imposible de determinar mediante el estudio de frecuencias de alelos. Los avances en el conocimiento del genoma y en el uso tanto del ADN mitocondrial como del cromosoma-y(en los que sigue trabajando Cavalli-Sforza), han permitido conocer esta cuantía.
Así parece quedar claro que la difusión démica fue más importante en el sur, centro y este europeos, que en el oeste y norte, aunque no de manera absoluta tampoco en las del sur, centro y este, teniendo que sumar a la difusión démica la difusión cultural, esta última más importante en zonas del norte y oeste.
Como conclusión, el modelo de difusión démica explica bastante bien, a excepción de Europa, la expansión de muchas poblaciones, siendo para Europa la combinación de difusión démica y cultural, la cual puede incluir transmisión de lenguas en el plano cultural(Cavalli-Sforza, Renfrew).
En una publicación de 1.995, basándose en una selección del genoma sometida a selección a partir de HLA, Arnaiz-Villena y colAboradores postularon un origen común de peninsulares y magrebíes.
Sin embargo estudios posteriores utilizando el cromosoma-y parecían contradecir esto (Elena Bosch y colAboradores en el 2.000, Comas y colAboradores el mismo año en otro estudio). Otros estudios han ido corroborando las marcadas diferencias entre estas poblaciones (Bosch y colAboradores en el 2.001), recientemente de nuevo corroborados por un nuevo estudio del hg E3b (“Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa”, 2.004). En todos estos estudios se confirma la diferencia entre las dos poblaciones, cada una descendiente de linajes masculinos diferentes. Esta diferenciación se inició durante la primera colonización de homo sapiens sapiens en tiempos del paleolítico superior tanto en uno como en otro lado y de la cual descienden la mayoría de los pobladores actuales respectivos: por un lado los ibéricos, afines al resto de los Europeos occidentales(hg R1b), venidos a través de Asia, y por otro los magrebíes, descendientes de su propio linaje africano caucasoide (hg E3b), venidos de algún lugar del noreste de África. Así se deduce que el estrecho de Gibraltar ha servido de barrera genética entre las dos poblaciones, al menos durante un tiempo en la que ambas crecieron lo suficiente por separado.
En cuanto a la razón de los diferentes resultados de Arnaiz-Villena, esta es debida a que al seleccionar una sola parte del genoma, la naturaleza aleatoria de la deriva genética y la acción de la selección pueden producir este tipo de desviaciones; para evitarlas, la interpretación debe apoyarse en la información conjunta del máximo número posible de genes y no en una sola región del genoma.
Sin embargo, volviendo a los estudios de Bosch, Comes..., a pesar que el estrecho de Gibraltar a servido de barrera para el intercambio de genes, tanto Iberia como el norte de África han recibido flujos de pequeña magnitud hacia uno y otro lado.
Curiosamente cuando se comparan los datos del ADN mitocondrial con los del cromosoma-y, resulta que los linajes subsaharianos heredados por vía materna se hallan a una frecuencia más elevada en magrebíes e ibéricos que los linajes paternos, lo que indicaría una diferencia entre sexos en la movilidad de los individuos desde el sur del desierto del Sahara. Esta observación genética debe contrastarse con datos sociales de movilidad y comercio de esclavos. Además de eso, las poblaciones femeninas en general presentan mayor movilidad que las masculinas; Cavalli-Sforza interpreta esta movilidad como que las mujeres han tendido históricamente (y prehistóricamente)a desplazarse al lugar de origen de su compañero.
COMPOSICIÓN DESDE LA PERSPECTIVA DEL ADN MITOCONDRIAL.
Hay que decir que los porcentajes dados a continuación son orientativos, y difieren ligeramente de una publicación a otra, pero en todas ofrecen porcentajes similares. Unos son según los datos publicados en el 2.003 sobre las poblaciones del oeste del mediterráneo(University College London), otros sobre un estudio sobre pasiegos que incluían otras poblaciones ibéricas, también en el 2.003.
H y V tienen un origen en el norte peninsular y representan los haplogrupos más extendidos en Iberia, siendo H el haplogrupo más extendido en el oeste europeo, con un 49.2% de media en Iberia. Hg V alcanza un 10% en vascos, 21.6% en cantabros, y 5.2% en gallegos, pero es muy baja en el centro peninsular y Valencia, lo cual indica que la mutación de V es reciente, poco antes de la recolonización postglaciar, hace unos 17.000 años; en el norte de África está presente en saharauis(17.9%) y bereberes del sur (10%); la media en norte africanos es del 6,8%. H también alcanza un porcentaje importante en todas las poblaciones del norte de África, con un 25.6% en argelinos, 26,5% en tunicios y un 36.8% en bereberes marroquíes; esta alta frecuencia entre norteafricanos se explica mediante una expansión neolítica desde Iberia (que coincide con la de la cultura del vaso campaniforme, originada en Iberia durante el eneolítico).
J y T presentan sus frecuencias más altas en Italia con valores del 15%. Iberia muestra una distribución más heterogénea con valores del 6,6% en valencianos y de 18,7% en el sur de Portugal. El noroeste de África muestra una distribución similar de J y T que los europeos, siendo más altos en saharauis y mauritanos, estando ausente más al sur. J se subdivide en dos: J1 y J2; J1 se cree que vino a través de una ruta continental, mientras que J2 lo hizo a través de la costa mediterránea.
Hg U6(se corresponde con el E3b2*), se originó probablemente en Oriente Medio y extendió al noroeste de Africa durante el paleolítico superior hace unos 30.000 años(con altas frecuencias en mozabitas, comunidad de origen bereber que vive en Argelia(28,2%) y mauritanos(20%), disminuyendo en tunicios(4,2%) y arabes marroquíes(8%), estando ausente en argelinos). Tiene una distribución poco densa en Iberia. Está presente mayormente en la zona sudoeste peninsular, pero a bajas frecuencias, con menos del 7%, alcanzando una media del 1.8%. Está ausente en vascos, catalanes, valencianos, centro y sur de Portugal. En las Islas Canarias alcanza el 14%, debido a la clara herencia guanche. En Italia U6 sólo está presente en Sicilia( 1%). U6 se subdivide en U6a y U6b, ambos presentes en Iberia y Canarias, pero con ausencia de U6b en el norte de África, lo cual indica un movimiento de pequeños grupos hacia la península en épocas prehistóricas; así, basado en datos arqueológicos y antropológicos se cree que estos pequeños grupos primitivos fueron empujados por los portadores de la cultura capsiense, pasando los primeros (huyendo de los segundos), a Canarias y la Península Ibérica; concretamente el U6b1 tiene una frecuencia del 13% en canarios, alcanzando 0.2% en la península, lo cual indica una reciente migración hacia la península efectuada por guanches.
El hg L(que incluye el L1, L2, L3) también está presente en Iberia, pero sólo alcanza un máximo del 6.8% en el centro de Portugal, siendo en el resto de la península muy escaso. La presencia de este linaje se explica por el comercio de esclavos durante la época colonialista europea.
El resto de haplogrupos presentes, según los datos del estudio sobre las poblaciones del oeste del meditarráneo del 2.003, donde da datos de las poblaciones de Valencia, Cataluña, País Vasco, centro peninsular, Portugal, Andalucía y Galicia, van como siguen. En conjunto U, sin contar la variante U6, el centro peninsular sería el de más porcentaje con un 20%, disminuyendo hacia la periferia, con un mínimo del 9% en catalanes. K alcanza un máximo del 10% en valencianos siendo en conjunto más abundante en la periferia que en el centro peninsular, donde alcanza el mínimo de 2%; en Galicia también es bajo (3,9%). I sólo está representado en Andalucía, con un 1,9% y Portugal, con frecuencias similares. W presenta una irregular dispersión; alcanza el 5% en Cataluña, está ausente en Valencia y País Vasco y entre 1 y 2% en el resto. Por último X, con bajas frecuencias en todas las poblaciones, con un pico en el centro de Portugal del 3,6%.
COMPOSICIÓN DESDE LA PERSPECTIVA DEL CROMOSOMA-Y.
En julio del 2.004, se publicó un documento que hacía un estudio desde la perspectiva del cromosoma-y sobre la Península Ibérica. En él se comparaban 11 poblaciones ibéricas(Sevilla, Huelva, Cadiz, Córdoba, Malaga, N. Portugal, León, Galicia, Cantabria, Valencia y Castilla) con las vascas y catalanas para tener una visión más clara de la aportación genética vía paterna en la península y de paso averiguar qué procesos demográficos habían incidido en su estructura genética. Los resultados revelaron una reducida variación interpoblacional del cromosoma-y (1,2%), que apunta a una limitada heterogeneidad en la región. También se observó que ni una vieja o reciente expansión desde Oriente Medio ni del norte de África a influido en la diversidad actual del cromosoma-y en Iberia(también corroborado por restos arqueológicos), por lo que los patrones geográficos pueden ser identificados.
La población que más atención a recibido en estudios genéticos es la vasca. La creencia de que su cultura y lengua tiene un origen ancestral, ha hecho que gocen de la máxima atención de los genetistas. Y no es para menos; su composición genética presenta la proporción más alta del haplogrupo R1b con un 89%(que coincide con la expansión del hg H del ADN mitocondrial, igualmente alto en vascos) y a penas posee en su composición genética haplogrupos de origen neolítico-anatolio(J, G, E3b). La composición genética también demuestra un cierto aislamiento y endogamia. Sin embargo no es una población aislada y hoy en día queda demostrado su relación con sus poblaciones vecinas ibéricas.
Los pasiegos es un grupo peculiar dentro de las poblaciones ibéricas, ya que recientemente se ha revelado un alto porcentaje del hg E3b de origen norteafricano con un 41% de individuos pertenecientes a este linaje (“Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa”, 2.004), demostrando ser una población muy heterogénea compuesta por grupos de diferente origen; uno peninsular y otro bereber seguramente venidos durante la invasión musulmana.
El resto de poblaciones, a excepción de la catalana, muestra una composición más heterogénea, fruto de las sucesivas invasiones recibidas a lo largo de la historia de la Península, aunque las aportaciones de estos linajes son mucho más modestas, pues la población ibérica ya era abundante desde el neolítico y estos eran a su vez los descendientes de los primeros pobladores portadores de la cultura auriñaciense.
Según este estudio el 61,4% pertenecen al linaje más antiguo, el denominado R1b, el más común en el oeste europeo y que define en más del 50% de los cromosomas-y ibéricos, siendo el más frecuente en todas las poblaciones.
Los linajes de origen africano alcanzan un 10,8% del total, alcanzando en algunas poblaciones altas frecuencias como es el caso de Galicia con un 46% y Malaga con un 30.6%. Curiosamente en el resto de poblaciones andaluzas el porcentaje no es muy alto, entre un 3,6% en Cadiz y un 13,5% en Huelva. N.Portugal 15.5%, León 10%, Valencia 16,2% Cantabria 12,9%, Castilla 4,8%, en catalanes no se ha registrado ningún linaje norteafricano y en vascos un 2,2%. Están relacionados por la llegada de grupos norteafricanos durante la invasión musulmana.
Por otra parte los haplogrupos I alcanzan altas proporciones en Andalucia y levante, aunque es bastante alto en Castilla (33,3%)y comparable a los porcentajes de Cerdeña, donde el porcentaje es el más alto de toda Europa. El subhaplogrupo I1b2 está relacionado con un grupo que presumiblemente usó Iberia como refugio durante la última glaciación, aunque su aporte a estas poblaciones es pequeño. También tiene un porcentaje en vascos del 8,8%.
El haplogrupo J tiene una media del 6,2%. Muestra una tendencia decreciente desde el sudoeste hacia Cataluña, con 17,9% en Andalucía, alrededor del 5% en León, Galicia, y Cantabria, 9,7% en Valencia siendo casi inexistente en Cataluña y País vasco. Está relacionado con la expansión neolítica originaria de Oriente Medio.
El grupo G no es considerado importante, por su baja frecuencia en las poblaciones, aunque está relacionado también con la expansión neolítica. Tiene un porcentaje entre el 6 y el 8% en N. Portugal, León, Cantabria y Cataluña, mientras que en el sur está en muy bajas proporciones y es inexistente en vascos, gallegos, castellanos y valencianos.
El estudio concluye resaltando la poca distancia genética entre las poblaciones ibéricas, incluida la vasca, una vez se quitan los datos de aportes genéticos minoritarios, que son las que crean cierta distancia genética.
A pesar de la compleja historia que ha sufrido la Península Ibérica el estudio demuestra la importante aportación de las poblaciones paleolíticas, la reducida estructura genética entre las poblaciones ibéricas y, por tanto, la gran afinidad de todas ellas entre sí, genéticamente hablando.
FENOTIPOS.
El primer genetista del mundo fue Mendel. Sus descubrimientos le llevaron a postular las tres leyes que llevan su nombre pudiendo así conocer cómo se producía la herencia genética. Así, él demostró que los caracteres se transmitían independientemente a través de unidades hereditarias que eran por naturaleza discontinuas; además estos caracteres no se fundían para siempre en la siguiente generación sino que se segregaban en la nueva formación de los gametos(espermatozoide y óvulo), es decir, un modelo de herencia no mezclada. Cuando el individuo produce sus células sexuales en estos sólo viajan la mitad de sus genes, mezclandose con la otra mitad del otro progenitor durante la fecundación. El nuevo individuo resulta así con una dotación genética doble cuyos caracteres hereditarios se regulaban mediante alelos dominantes y recesivos.
Luego vino la teoría de la mutación de Hugo de Vries, cambio espontáneo y al azar el cual era un cambio genético de envergadura, creando nuevas especies si las mutaciones eran las apropiadas para sobrevivir en el medio.
Si se suma la adaptación al medio mediante la selección natural, ya tenemos una idea de cómo se producen cambios en la morfología de los individuos y la diferenciación de unos grupos de otros.
Entonces además de depender de nuestra herencia genética, dependemos de mutaciones que se añaden a nuestro acerbo genético y que deben ser aptas para permitirnos la vida en un determinado medio. Además nuestra morfología dependerá de dietas, enfermedades, que afectarán a nuestra salud y desarrollo individual.
Se podría pensar que los diferentes haplogrupos coinciden con las razas actuales. Pero el problema es que las mutaciones son aleatorias tanto para la mutación en si como para el tiempo en el que se produce, por lo que dos poblaciones con fenotipos diferentes pueden pertenecer al mismo haplogrupo y parecer de mundos diferentes. En esto hay muchos factores que intervienen, como ya se ha dicho arriba.
Un ejemplo claro de esto es el nuestro: Los europeos somos descendientes de nuestros antepasados paleolíticos, fundamentalmente de los portadores de la cultura auriñaciense a Europa, cuyo representante en el registro fósil es el hombre de cromañon. Éste era de elevada estatura, entre 1,70 y 1,85 m, caderas estrechas, dolicocéfalos y de cara ancha.
Pero durante la última glaciación vemos una evolución de éste tipo, viendo como tiende a ser más bajo. Así, observando el registro fósil, tenemos al hombre de Combe-Capelle, correspondiente a la cultura perigoriense, de 1,63m y dolicocéfalo, ya empieza a mostrar rasgos de la raza mediterránea que tendrá su difusión en tiempos del mesolítico y neolítico por toda Europa. Lo mismo se puede afirmar sobre Los hombres de Chacelade, que se consideran descendientes del de Combe-Capelle.
La razón de esto se cree que se debe a diferentes factores:
Una adaptación a un ambiente extremadamente frío, en donde los individuos “redondos” o menos alargados, mantienen el calor mejor que los individuos “alargados” o altos. Esto no implica una menor robustez, siendo las poblaciones más bajas pero bien construidas.
Para el antropólogo Lionel Sims la pérdida de robustez se debe a una etapa en la que desaparecieron los grandes animales, cuya caza implicaba un trabajo en equipo, camuflaje y experiencia, con una vida nómada, sin enfermedades típicas de las poblaciones sedentarias, hacía de esta una vida más excitante y sana; al depender más de la caza de los pequeños animales, más difíciles de cazar, y al verse envueltos en un rápido cambio en su ambiente, el paso fue más bien traumático; al principio el paso a una vida sedentaria al adoptar la agricultura resultaría aburrida para individuos acostumbrados a una actividad más excitante; eso sumado al desarrollo de enfermedades típicas del sedentarismo y desconocidas para nuestros antepasados hasta entonces, contribuyó a un empeoramiento de la salud y, por tanto, dando lugar a individuos con esqueletos más gráciles, afectando también a la altura.
El color muy claro de los pobladores de las regiones del norte de Europa se debe, según Marvin Harris, a que pueblos de fenotipo mediterráneo producían alimentos agrícolas, derivados lácteos, y obtenían la vitamina D del sol. Pero al comenzar su lenta ascensión hacia el norte, sufrieron un déficit de vitamina D (que se obtiene normalmente del pescado marino) y de calcio que hizo que la selección natural favoreciese la piel blanca, por su mayor receptividad frente a las radiaciones solares, y también por su mayor capacidad para producir la enzima llamada lactasa que posibilita la ingestión de grandes cantidades de leche fresca, alimento básico en los territorios nórdicos. Así, la supervivencia de hijos de piel clara, más fuertes y más sanos por lo general que los hijos morenos en ambientes de poca radiación solar con déficits alimenticios, unido a la alta mortalidad infantil de esas épocas, dio como resultado a una mayor proporción de rubios en las poblaciones nórdicas. Para Harris la mayor proporción del tipo nórdico se produjo hace 6.000 años, pero en realidad se puede asegurar que entre las poblaciones paleolíticas (caucasoides o no) ya habían individuos rubios, aunque no fueran ni mucho menos mayoritarios con respecto a los morenos.
Para Henri V. Vallois, los nórdicos son "un conjunto heterogéneo que corresponde a la despigmentación independiente de grupos diferentes". Este punto de vista concuerda con la diversa composición genética que presentan las poblaciones del norte europeo, y que sin embargo son la máxima representación del tipo nórdico.
Viendo la variedad de fenotipos tan grande dentro de un mismo haplogrupo nos damos cuenta que los cambios morfológicos suelen producirse más rápido que las mutaciones que dan lugar a nuevos haplogrupos, por lo que no se puede relacionar haplogrupo con fenotipo.
LENGUAS.
Cavalli-Sforza cree que los linajes del cromosoma-y pueden estar asociados con los mayores grupos lingüísticos del planeta. Y la verdad es que viendo la distribución de las principales familias lingüísticas del mundo y comparandolas con los haplogrupos correspondientes, vemos que lo que era imposible con los fenotipos, no lo es tanto con las lenguas. Aquí si que vemos cierta correlación para las lenguas afroasiáticas, urálicas, amerindias(antes de la llegada de europeos), etc, aunque con excepciones como la del grupo indoeuropeo, en la que hay controversia sobre sus orígenes.
Los hablantes de las lenguas indoeuropeas abarcan grupos humanos de origen lo suficientemente separado en el tiempo para que todos hablaran unas lenguas tan parecidas entre sí que se pudieran incluir en la misma familia. Así tenemos a nuestros ancestros que habitaban el occidente europeo antes de la última glaciación, y los que habitaban el este. Ambos recolonizaron Europa después de la retirada de los hielos y ambos utilizaban su propio lenguaje. Una separación de más de 20.000 años no puede dar lugar a grupos que hablaran lenguajes parecidos que se conservaran igual durante tal espacio de tiempo. Por lo que la única alternativa que nos queda es la de una sustitución de unas lenguas por otras.
Tanto el genetista Cavalli-Sforza como el arqueólogo Colin Renfrew(siguiendo el modelo de ola en avance, basandose en el registro arqueológico y ahora también en el genético), creen que las lenguas indoeuropeas fueron traídas por los primeros grupos neolíticos, portadores de la agricultura y la ganadería desde su ubicación original, en Anatolia.
El filólogo finlandés Kalevi Wiik se ha basado en estos últimos descubrimientos genéticos para lanzar su teoría según la cual entre hace 23.000 y 8.000 años Europa estaba dividida en tres regiones; dos llamada Ba(protovascas) al oeste, y U (uralicas) al noreste, poblada por cazadores de grandes animales que abundaban en esas épocas. Una tercera zona denominada X que corresponde a la zona centro y sur de Europa y que estaba poblada, según él, por cazadores de pequeños animales, zona que estaba fragmentada en varias lenguas que no han sobrevivido.

Entonces hace unos 5.500 años, hubo un cambio radical que hizo que las zonas Ba y U entraran en recesión; este fue la desaparición de los grandes animales en esas zonas, mientras que en la zona X empezaron a adoptar las formas de viva Neolítica, domesticando animales y desarrollando la agricultura, que les hizo más fuertes económicamente a la vez que desarrollaron sistemas de organización complejas, creciendo en número. Es aquí donde Kalevi Wiik defiende que desde los Balcanes y Grecia ( los exportadores del Neolítico, según él) se expandió el indoeuropeo, sirviendo como lengua vehicular para los habitantes de la zona X, reemplazando las lenguas de la zona X y convirtiendo más tarde a los menos exitosos habitantes de la zona Ba y U.

Finalmente, justo antes de la conquista romana, en las zonas más occidentales, todavía habrían muchos hablantes de lenguas Ba que habrían adoptado la agricultura y ganadería sin perder sus lenguas, aunque con gran influencia indoeuropea, como la de los ligures, las lenguas ibéricas y/o euskéricas, de los pictos o cruithin en Escocia e Irlanda, y de los pridyn en Gales,

hasta que finalmente desaparecieron por conquista de romanos o gaélicos, con la excepción del Vasco.

Para algunos lingüistas, entre los que se encuentran Theo Vennemann, la lengua vasca es la última representante de una familia de lenguas extendidas por todo el occidente europeo en épocas antiguas y que fueron sustituidas por las lenguas indoeuropeas, portadas por grupos originariamente venidos de Anatolia y sudeste europeo.
Bibliografía.
LAboratorio de Evolución, Facultad de Ciencias, Universidad de la República(Uruguay):
http://evolucion.fcien.edu.uy/Diapositivas/GenesHumanos4d.pdf
De la web de Cristina Díaz. Apartado “de dónde venimos”(La superexplosión del Toba): http://www.mundofree.com/origenes/sapiens/donde.htm
Glosario de Términos Genéticos: http://www.genome.gov/sglossary.cfm
Red Cívica Americana. El Árbol genealógico de la Familias humana:
http://www.redcivica.com.uy/archivos/html/revista/tomoIII/El%20arbol%20genealogico.htm
Tesis de Stèphanie Plaza. Anàlisi de la diversitat del genoma mitocondrial en poblacions humanes:
http://www.tdx.cesca.es/TESIS_UPF/AVAILABLE/TDX-0709104-123956//tspp1de1.pdf
Técnicas de análisis del ADN en genética forense: ANÁLISIS DEL ADN EN GENÉTICA FORENSE
http://members.fortunecity.es/robertexto/archivo7/adn_forense.htm
Patología y mecanismos de los trastornos genéticos:
http://ventanadelavida.jesussaveus.com/apuntesmedicina/genetica.htm
Orígenes de los Humanos Modernos: ¿Multiregional o Fuera de África?
Por Donald Johanson:
http://www.actionbioscience.org/esp/evolution/johanson.html
Juan Luis Arsuaga-EVOLUCIÓN HUMANA-C.N.I.C.E.:
http://www.cnice.mecd.es/tematicas/evolucion/2002_07/2002_07_01.html
Tesis de Siiri Rootsi: HUMAN Y-CHROMOSOMAL VARIATION IN EUROPEAN POPULATION:
http://www.tymri.ut.ee/PhD/2004/SRootsi_thesis.pdf
British teacher finds long-lost relative: 9,000-year-old man
http://www.standardtimes.com/daily/03-97/03-09-97/a09wn056.htm
Oetzi, el hombre de hielo http://news.bbc.co.uk/hi/spanish/science/newsid_3231000/3231879.stm
Las siete hijas de Eva.
Bryan Sykes http://usbiver.tripod.com/hija.html
mtDNA haplogroup of L. David Roper
http://www.roperld.com/mtDNA.htm
The Human Family Tree: 10 Adams and 18 Eves By Nicholas Wade
http://www.ramsdale.org/dna7.htm
World haplogoups maps, J. D. Mcdonald http://www.scs.uiuc.edu/~mcdonald/WorldHaplogroupsMaps.pdf
Inferring Human History: Clues from
Y-Chromosome Haplotypes
P.A. UNDERHILL http://hpgl.stanford.edu/publications/Underhill_2004_p487-494.pdf
The Molecular Dissection of mtDNA Haplogroup H Confirms That the
Franco-Cantabrian Glacial Refuge Was a Major Source for the European
Gene Pool
Alessandro Achilli y colAboradores, http://evolutsioon.ut.ee/publications/Achilli2004.pdf
Demic Diffusion as the Basic Process of Human Expansion. http://hpgl.stanford.edu/publications/Examining_2003_chapter7.pdf
Expansión de mtDNA mundial según J. D. Mcdonald. http://www.mcdonald.cam.ac.uk/genetics/mtDNAworld/one.html
Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa, 2.004. http://www.familytreedna.com/pdf/hape3b.pdf
Reduced genetic structure of the Iberian peninsula
revealed by Y-chromosome analysis: implications for
population demography. http://hpgl.stanford.edu/publications/EJHG_2004_v12_p855.pdf
A Signal, from Human mtDNA, of Postglacial Recolonization in Europe. http://www.oxfordancestors.com/papers/mtDNA01%20Postglacial.pdf
mtDNA Analysis in Ancient Basque Populations http://www.journals.uchicago.edu/AJHG/ journal/issues/v69n4/012962/012962.web.pdf
Alu insertion polymorphisms in NW Africa and the Iberian Peninsula:
evidence for a strong genetic boundary through the Gibraltar Straits.
http://batzerlab.lsu.edu/Publications/ Comas%20et%20al.%202000%20Hum%20Genet.pdf
Mitochondrial DNA transit between West Asia and North Africa
inferred from U6 phylogeography. http://www.biomedcentral.com/content/pdf/1471-2156-4-15.pdf
The Genetic Legacy of
Paleolithic Homo sapiens
sapiens in Extant Europeans: A
Y Chromosome Perspective. http://hpgl.stanford.edu/publications/ Science_2000_v290_p1155.pdf
El enigma de la esfinge, Juan Luís Arsuaga.
Enciclopedia Universal Multimedia ©Micronet S.A. 1999/2000.




















Comentarios
 Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Pulsa este icono si opinas que la información está fuera de lugar, no tiene rigor o es de nulo interés.
Tu único clic no la borarrá, pero contribuirá a que la sabiduría del grupo pueda funcionar correctamente.
Si te registras como usuario, podrás añadir comentarios a este artículo.
Microsatélites y la expansión de los seres humanos modernos
Existen en la actualidad dos hipótesis predominantes para explicar el origen y dispersión mundial de los seres humanos anatómicamente modernos; esto es, todas las formas de Homo sapiens que presentan las características craneofaciales y esqueléticas del hombre actual. Una de ellas, denominada multirregionalista, postula que los seres humanos han tenido múltiples orígenes independientes a partir de un cierto nivel del proceso hacia la hominización, por el cual se describe la evolución de las formas fósiles más antiguas hasta el hombre actual. Los puntos de origen se ubican fundamentalmente en Europa, África y el este asiático, y son representados por formas de Homo erectus (Pitecantropos, Sinantropos, etc.). Este punto de vista es liderado, entre otros, por Milford Wolpoff de la Universidad de Michigan. La otra hipótesis postula un origen único para Homo sapiens que, si bien refleja una concepción clásica de la unidad de origen de la especie humana, en los últimos tiempos ha sido fortalecida por el avance de las técnicas genéticas. Estas han permitido calcular que el hombre actual que puebla todos los continentes desciende de una única población –aún no descubierta– cuyos ancestros debieron habitar una pequeña porción de territorio al sur del África subsahariana. Dado que la teoría sostiene la existencia de un único centro de origen africano con posteriores migraciones, se la ha denominado out of Africa. En su forma extrema, se complementa con el modelo de Migración-Reemplazo (MMR), según el cual los hombres anatómicamente modernos, en su migración hacia el norte, este y oeste, eliminaron de la faz de la Tierra a todas las formas homínidas preexistentes, de modo que no hubo posibilidad –según el razonamiento– de mezcla racial alguna.
La gran similitud morfológica entre las poblaciones humanas modernas y su diferenciación respecto de cualquier otra especie homínida, contribuye a sostener el criterio de origen único, lo cual es consistente con los resultados de los análisis genéticos. Según autores como Takahata, el 90 por ciento de las secuencias de ADN estudiadas en Homo sapiens indican un origen africano del antecesor común más reciente. Uno de los principales soportes del MMR es que la variación del ácido desoxirribonucleico mitocondrial (ADNmt) de los hombres de Neanderthal cae fuera del rango de variación de los seres humanos modernos, sugiriendo que no efectuaron contribución genética alguna. Sin embargo, no está todo dicho al respecto ya que varios autores han encontrado superposición en las distribuciones de variación genética en ambas formas homínidas.
Los últimos avances de la investigación parecen consolidar la hipótesis out of Africa. Un estudio realizado por investigadores de la Universidad de Stanford y de la Academia de Ciencias de Rusia, bajo la coordinación del profesor MW Feldman, demuestra que los microsatélites –pequeños segmentos de ADN repetitivo caracterizados por una alta tasa de mutación– en virtud de su transmisión generacional se convierten en una excelente herramienta para estimar el momento en que dos poblaciones divergen. Estos investigadores compararon 377 marcadores de microsatélites en ADN, tomados de 1056 individuos procedentes de 52 regiones esparcidas por el mundo. El análisis reveló una estrecha afinidad entre dos poblaciones de cazadores-recolectores de África subsahariana: los pigmeos mbuti de la cuenca del Congo y los bosquimanos procedentes de Boswana y Namibia. Estas dos poblaciones tienen un modo de subsistencia típico de caza-recolección. Esta forma de vida, basada en la caza de pequeños animales y en la recolección de vegetales silvestres comestibles, es considerada como la más antigua –si bien aún hoy hay sociedades que la siguen practicando– y contribuye a apoyar la idea de que estos grupos de pigmeos y bosquimanos descienden de los primeros ancestros que alcanzaron el nivel evolutivo correspondiente a los seres humanos anatómicamente modernos. Se ha estimado que esta población hipotética era muy pequeña –unos 2000 individuos– y habría comenzado sus primeros movimientos migratorios hace unos 70.000 años (71.142 años para ser exacto). Los resultados concuerdan con los 66.000 años obtenidos en un estudio previo realizado por Feldman y colAboradores, obtenidos sobre la base del análisis en cromosoma Y de más de 1000 hombres procedentes de 21 poblaciones diferentes. Según Feldman, una vez fuera de África la expansión continuó por Eurasia, Oceanía, este de Asia y, finalmente, América.
Algo llamativo, pero que concuerda con lo visto hasta el momento, es que mientras se desarrollaban estos acontecimientos, el resto de África no estaba inactivo. Los resultados sugieren que también hubo en la región un conjunto de poblaciones que practicaban la agricultura luego de separarse de los cazadores recolectores en una fecha que todavía es por demás imprecisa. Esto permite conjeturar con un alto grado de probabilidad que hubo dos procesos expansivos, ambos originados en África. Uno, el de los cazadores-recolectores que migraban porque su tipo de actividad impedía el crecimiento poblacional más allá de los límites indicados (unos pocos miles de individuos), y otro posterior de los agricultores que, gracias a su economía, incrementaron enormemente la densidad poblacional a varios millones y ocuparon diversas regiones de Asia.
Una de las consecuencias más importantes del efecto out of Africa es que, por ser la densidad de los cazadores-recolectores normalmente tan baja, la población que hubiere dado origen a toda la humanidad actual debía tener forzosamente una variabilidad genética prácticamente ínfima. Sobre todo cuando se la compara, por ejemplo, con uno de los parientes actuales más próximos al hombre. Un pequeño grupo de chimpancés puede tener más diversidad genética que los seis mil millones de humanos que viven en la actualidad. Si se piensa que todos los homínidos provienen de un antecesor común con los chimpancés hace unos cinco millones de años, se concluiría que el tiempo transcurrido fue más que suficiente para desarrollar cambios genéticos sustanciales. Sin embargo, la ausencia de estos cambios –sugerida por la similitud genética actual entre los géneros Pan y Homo– indica que el pool génico humano se redujo sensiblemente en el pasado reciente desde hace unos 100.000 años y, de este modo, hubo una limitación a la variación genética de las poblaciones actuales. Esta estimación no se modifica ni aun admitiendo que no una sino varias de estas poblaciones podrían haber existido en el pasado remoto, pues habrían estado genéticamente aisladas entre sí y, en consecuencia, solo unas pocas pudieron dar origen a los hombres actuales. La tendencia a la ´uniformidad genética´ debió agravarse por más de un efecto ´cuello de botella´ –una restricción de la variabilidad genética provocada por barreras geográficas o ecológicas– que, sin duda, han ocurrido en el transcurso de tan extenso proceso migratorio.
Por último, podemos ocuparnos de un hecho paralelo al estudio en sí pero no por ello menos relevante y que se refiere al comportamiento de los individuos empleados en el estudio. Se afirma que el análisis de ADN confirmó la respuesta de la mayoría de los 1056 participantes respecto de sus ancestros, determinando que dicho procedimiento constituye una vía útil y válida de información. Esto viene a colación debido a que algunos miembros de la comunidad médica argumentan que el profesional no debe interrogar a los pacientes sobre sus ancestros, porque carece de rigor; sin embargo se halló que la ancestría informada fue consistente con los resultados del estudio. Según Feldman y colAboradores, la ancestría declarada por los pacientes permite obtener información rápida sobre factores ambientales, diferencias geográficas y culturales y comportamientos diversos que, además de sus implicancias teóricas, pueden ser importantes factores de riesgo para ciertos tipos de enfermedad.
Héctor M Pucciarelli
Doctor en Ciencias Naturales, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Profesor Titular Ordinario de Antropología Biológica I, Facultad de Ciencias Naturales y Museo, UNLP. Investigador Superior del CONICET. Jefe de la División Antropología del Museo de La Plata, FCNyM, UNLP.
http://www.ciencia-hoy.retina.ar/hoy82/microsatelite.htm
Antes de lanzarse a defender que los "humanos morfologicamente modernos" no descienden de los neandertales porque ambos poseen adns mitocondriales diferentes es mejor informarse.
Ningún científico afirma nada parecido a lo anterior. Lo máximo que se puede decir es que "las secuenciaciones llevadas a cAbo apoyan la idea de que no hay contribución de los neandertales al pool de los genes mitocondriales de los humanos modernos. Sin embargo esto no excluye la posibilidad de que los neandertales hayan contribuido genéticamente con un aporte de adn nuclear" (Hoss M: Neanderthal population genetics, Nature, 2000, vol. 404, 30 March, p. 453-4).
Mucho cuidado con extraer conclusiones precipitadas.
Hola Onnega.
La verdad que lo de la idea de que los neandertales se extinguieron sin dejar descendientes en los humanos actuales no es una idea nueva de la genética sino que es una vieja discusión de paleontólogos antropólogos, etc.
Comparando fósiles de neandertal con humanos modernos, el primero prensenta importantes diferencias con el segundo y con el europeo en particular (por ser este el que invadió el territorio del neandertal, que fue principalmente Europa).
Su cerebro era más grande que el nuestro por término medio; su cara era más grande, con un arco supraorbital muy marcado y una nariz muy robusta; su maxilar inferior era también robusto y el mentón achatado. Los dientes eran mucho más grandes y estaban dispuestos en una curva en forma de U, no en forma de parábola como los nuestros. Su cabeza era sustentada por músculos cortos y muy robustos en la nuca. Era muy bajo (1,60m) y mucho más musculoso que el cromañon.
El hombre de neandertal evolucionó entre hace 200.000 y 100.000 años( glaciación de Riss e interglacial de Riss-Würm) en Europa y se le asocia a la cultura musteriense. Nuestro antepasado el hombre de cromañon hizo su aparición de repente en Europa hace 40.000 años(glaciación de Würm), portando la cultura auriñaciense, por lo que hace imposible ver pasos intermedios entre las dos especies que las pueda relacionar. Lo que si se cree es que ambos descendían del homo heidelbergensis y el homo antecessor.
Además, como ya comenté en el artículo, los paleontólogos, genetistas, etc, aunque no descartan por completo una hibridación, nuevas pruebas apuntan en la dirección opuesta.
En el recientemente pasado año 2.004 se publicó un artículo en la revista británica nature en el que se comentaba que el neandertal llegaba a adulto a los 15 años, mientras que el hombre moderno finaliza su desarrollo entre los 18 y 21 años.
A esta conclusión se llegaba estudiando los dientes los cuales marcan las huellas del desarrollo del individuo. En el homo antecessor, homo heidelbergensis, hombre de cromañon y el hombre moderno el crecimiento del esmalte dental, después de la formación de la mitad superior del diente, es lento, por ello las estrías están muy próximas. En los neandertales estas estrias están bien separadas, lo que indica un crecimiento rápido.
El por qué de éste desarrolo acelerado podría deberse a la alta mortalidad que sufrían sus poblaciones debido al extremo ambiente ostil en el que vivían. Otra explicación es que consumían muchas calorías y poseían un metAbolismo acelerado.
De cualquier manera este descubrimiento nos aleja, según los científicos, de nuestro primo el neandertal.
http://www.portaldehistoria.com/secciones/portada2/news/news_item.asp?NewsID=89
Aquí hay un enlace donde se da cuenta de la primera extracción de ADN de Neandertal en España.
Se insiste enque no hubo intercambio genético con otras especies:
http://www.lne.es/secciones/noticia.jsp?pIdNoticia=259509&pIdSeccion=46&pNumEjemplar=821
"Los expertos, dirigidos por el paleobiólogo del Consejo Superior de Investigaciones Científicas (CSIC) Antonio Rosas, en colAboración con el investigador de la Universidad de Barcelona Carles Lalueza, han descubierto que los neandertales de la Península no eran genéticamente distintos a los del resto de Europa.
Datados hace 43.000 años, aportan además información genética suficiente para ratificar que no hubo intercambio genético con otras especies. Antonio Rosas, que ayer mostraba su satisfacción por lo que considera «un importante logro científico», se basa para descartar la hibridación en que el ADN obtenido de neandertales y cromañones es muy distinto: «Los genes que los hombres actuales comparten con los neandertales no son más parecidos que los que comparten con el chimpancé; por tanto no es significativo"
Saludos Onnega:
El ADN nuclear sí que se ha estudiado:
"La carrera por descifrar la secuencia completa del genoma humano y lograr la localización de los genes en los cromosomas fue ganada por la técnica automatizada del emprendimiento privado, que el 6 de abril de 2000 anunció el primer borrador"
http://www.biotech.bioetica.org/ap13.htm
En el ADN nuclear de cualquier célula de un individuo se encuentra toda la información de este.
La mitad de los genes en la fecundación la aporta el espermatozoide, la otra mitad el óvulo. Es dificil trazar la historia de poblaciones a partir de este ADN nuclear que se recombina en cada fecundación.
Partes del cromosoma-y no se recombina durante la fecundación, lo mismo que el ADN mitocondrial, y sólo mutan aleatoriamente por fallos de copia muy de vez en cuando. Es por ello por lo que sirve para el estudio de poblaciones.
Quizá el neandertal era de nuestra misma especie, es decir, no era un homínido diferenciado, pero todas las pruebas apuntan a que no se mezcló con nosotros.
No creo que los científicos del CSIC y la universidad de Barcelona sean tranposos, sólo aportan pruebas científicas. Sería muy interesante que los que creen que venimos del neandertal aportaran pruebas para defender su posición. La interpretación de los fósiles puede alimentar la controversia pero la genética es una ciencia exacta (o casi). Si se decantan claramente por la teoría del origen reciente es porque las pruebas genéticas son sólidas:
"Madrid, 14 de febrero, 2005.- Un equipo de investigadores españoles, dirigidos por el paleobiólogo del Consejo Superior de Investigaciones Científicas (CSIC) Antonio Rosas, en colAboración con el investigador de la Universidad de Barcelona Carles Lalueza, ha alcanzado un logro científico sin precedentes al conseguir extraer y secuenciar por primera vez ADN mitocondrial de restos neandertales en la Península Ibérica. Los fósiles analizados en el estudio proceden de la cueva asturiana de El Sidrón y tienen 43.000 años de antigüedad."
"El análisis de la secuencia de ADN obtenido ha permitido a los investigadores establecer que las distintas poblaciones de neandertales que habitaron Europa desde la Península Ibérica hasta el Cáucaso eran genéticamente similares. Esta conclusión, en contra de lo que algunas hipótesis previas hacían suponer, permite situar un origen común de los neandertales hace 250.000 años. Esta fecha coincide con la aparición de la morfología de neandertal clásico y concuerda asimismo con un posible evento de especiación para el origen de este Homo neanderthalensis."
http://www.csic.es/prensa/noticias2005/febrero05/14febrero2005.html
Otro sitio:
http://www.diariomedico.com/edicion/noticia/0,2458,594450,00.html
http://news.bbc.co.uk/hi/spanish/science/newsid_1470000/1470785.stm
BBC Mundo|Ciencia.
Jueves, 02 de agosto de 2001 - 17:20 GMT
Vecinos sí, ancestros no
Los neandertales tendrían sus características distintivas desde la infancia. (Reconstrucción de E. Daynès, París).
Un equipo suizo realizó una reconstrucción computarizada del hombre de Neandertal y concluyó que coexistió con la raza humana, pero nunca se mezcló.
Christoph Zollikofer y Marcia Ponce de León, de la Universidad de Zurich, crearon modelos en tres dimensiones a partir de cráneos de neandertales.
Las imágenes muestran características distintivas que estaban presentes desde la niñez, y quizás desde el vientre materno.
"Se trata de un fuerte argumento para la separación de las especies, pues significa que ambas se aislaron", dijo Zollikofer.
Las diferencias
El cráneo de un niño humano y de un niño neandertal.
El propósito del estudio era comparar el desarrollo desde la niñez de los primeros humanos y los hombres modernos.
Para esto, realizaron tomografías a 16 fósiles de individuos que vivieron en Europa, África y Asia entre 125.000 y 40.000 años atrás.
Según los investigadores, a la edad de dos años el hombre de Neandertal ya presentaba las características que lo distinguían del humano, como la quijada achatada y la frente inclinada.
Para ellos, esto sugiere que los neandertales eran una especie "hermana" del ser humano.
"No hemos visto ninguna evidencia de que haya habido una combinación de genes", dijo Zollikofer.
Misteriosa desaparición
Reconstrucciones virtuales desde los fósiles de un niño neandertal.
Los neandertales vivieron en Europa y Asia hace 100.000 años, y desaparecieron de la faz de la Tierra hace 30.000.
Su misteriosa desaparición ha sido sujeto de múltiples teorías y especulaciones.
Para algunos, estos hombres primitivos murieron porque una nueva raza humana acaparó los territorios en los que cazaban durante la última Era de Hielo.
Pero para otros, desaparecieron porque se mezclaron con la raza humana.
La investigación suiza negaría esta última teoría, junto con un estudio anterior que analizó el ADN de tres esqueletos de neandertales y concluyó que no eran nuestros ancestros sino una raza paralela a la humana.
Imágenes del LAboratorio de Multimedia de la Universidad de Zurich.
Gracias por tus aclaraciones sobre Oetzi, Giorgio, nadie mejor que tú para hacerlo; yo me he limitado a copiar la información de varios lugares. Concretamente donde dice lo de la frontera cerca de Alemania y Austria:
http://www.chs.ecu.edu.au/org/sebhs/hubiol/staff/chris%20meredith%20lectures/3.DNADetectives/5.mtDNAIceManNeander&7Eves.ppt
Sobre el encima llamado lactasa:
La intolerancia a la lactosa de la leche:
esta enfermedad es de origen genético y está muy ligada a grupos étnicos . Como ejemplo se puede poner a Dinamarca donde solamente la sufren un 2% de la población y en el otro extremo están Japón, China y los Aborígenes Australianos que la sufren casi un 100% en mayor o menor medida. Se cree que hay una relación directa entre la enzima lactasa que hidroliza la lactosa y la melanina responsable del color de piel. Cuanto mayor sea la concentración de melanina, menor es la de lactasa.
-------------------------------------------------------------------
La lactosa, un carbohidrato difícil de utilizar
La lactosa, que es un disacárido (galactosa+glucosa), es el azúcar de la leche. El organismo para su digestión y absorción tiene que desdoblarlo o hidrolizarlo para lo cual necesita de una enzima llamada lactasa. El organismo con la edad va disminuyendo la producción de lactasa, dependiendo de las razas. En la raza blanca, donde algunos pueblos han consumido leche tradicionalmente, la lactasa tiene una lenta disminución a partir de los 3 años. En cambio en razas como la negra, sólo hay lactasa hasta los 3 años, después desaparece.
Se dice que la razón por la que en determinadas razas blancas existe lactasa después de los 3 años es porque hay una relación directa entre la lactasa y la melanina responsable de la pigmentación de la piel. Los hombres que viven en zonas frías y tienen poca radiación solar, necesitan tener la piel más blanca para absorber la luz del sol, y de esa manera producir más vita mina D, necesaria para la absorción del calcio. La leche juega a favor de este mecanismo del calcio, y el organismo mantiene esta enzima para que sea posible su absorción y digestión.
Es posible, como se describe anteriormente, que la existencia de lactasa por encima de los 3 años se deba a la circunstancia del consumo de leche, pues no existe ningún alimento más que contenga lactosa.
Entre los consumidores de leche nos encontramos con personas que dicen digerir bien la leche, en realidad toleran la leche por mantener a estas edades suficiente producción de lactasa.
También nos encontramos otros muchos consumidores de leche y lácteos, que digieren mal la leche y otros lácteos, o que sufren de enfermedades como alergias, intolerancias, asma, problemas de piel, trastornos digestivos, etc., y que sus dolencias están directamente relacionadas con el consumo de leche y lácteos.
En estas personas que tienen un nivel bajo de lactasa, su intestino no puede hidrolizar la lactosa y ésta es fermentada por distintas bacterias, generando un efecto hiperósmico en las paredes intestinales causando meteorismo, inflamación e irritación intestinal, flatulencias, diarreas o estreñimiento, que no es otra cosa que la denominada intolerancia a la lactosa.
Esta fermentación anómala de la lactosa genera un efecto acidificador del organismo, reflejado en un aumento del nitrógeno en individuos con poca cantidad de lactasa. Además se ha comprobado que la reactividad ante las proteínas lácteas, antes descrita, aumenta con la intolerancia a la lactosa.
-------------------------------------------------------------
Como ves al final del artículo he puesto mucha bibliografía para que podáis investigar un poco sobre el tema, pero aquí:
http://hpgl.stanford.edu/publications.html
La Universidad de Stanford tiene publicados todos sus estudios, que se pueden descargar de manera gratuita.
Hola Aga:
Todas las razas caucásoides no son de piel blanca, como los indios de la India, y sin embargo antropológicamente sí son caucásicos y a demás están emparentados con el resto de europeos genéticamente;
Contrariamente a lo que ocurre con los Ainú(habitantes del Japón), que se consideran de raza caucasoide, y que se caracterizan por el color claro de la piel, por el color castaño claro de los ojos, que carecen de brida mongólica, por la pilosidad abundante, el cráneo fuertemente dolicocéfalo, la prominencia de los arcos superciliares, la estatura baja y el cuerpo no esbelto, pero sí muy robusto. Los Ainú están relacionados genéticamente con el resto de los asiáticos, sin embargo el fenotipo recuerda al de los europeos.
La cultura Inuit (esquimal) tradicional muestra la total adaptación a un entorno extremadamente frío, bloqueado por la nieve y el hielo, en el que los alimentos vegetales son casi inexistentes, los árboles escasos y el caribú, la foca, la morsa, la carne y la grasa de la ballena y el pescado las principales fuentes de alimento. La precariedad de este hábitat les ha llevado a desarrollar sorprendentes mecanismos de adaptación en todos los niveles. La naturaleza, por ejemplo, les ha provisto de un mecanismo de mantenimiento del calor que les permite resistir, mediante cambios en su metAbolismo, las temperaturas más extremas, la piel morena les proteje de la alta radiación solar debida al reflejo casi completo de la luz en la nieve. Su propia lengua refleja la importancia que los recursos de supervivencia adquieren en esta sociedad; así, por ejemplo, su vocabulario incluye una gran cantidad de palabras que pueden traducirse simplemente por "blanco", término tan amplio para ellos como podría ser el término "color" para nosotros. Para los esquimales es fundamental distinguir entre numerosos tipos de color blanco, puesto que de su mayor o menor brillo, de su distinta tonalidad o matiz (inapreciable para los que no viven entre hielos) depende en muchas ocasiones el poder distinguir que el piso está sólido y que se puede caminar por él, o que ciertos bloques de nieve son los idóneos para construir una vivienda, o que el tiempo es propicio para la caza. Los individuos de esta cultura aprendieron a utilizar los únicos recursos naturales de su mundo, como la nieve, el hielo, la piedra, la madera arrastrada por las aguas y unos cuantos animales, para vivir confortablemente y casi siempre con prosperidad.
Los individuos de este tronco racial se caracterizan por combinar rasgos físico-morfológicos típicos tanto de los pueblos xantodermos o amarillos como de los leucodermos o blancos, hasta el extremo de que algunos antropólogos los han considerado descendientes del grupo racial blanco mediterráneo. Genéticamente están relacionados con asiáticos, pero los europeos han contribuido de forma minoritaria en la composición genética.
en algunas poblaciones australianas, de piel oscura, hay niños rubios.
sacado de:
Enciclopedia Universal Multimedia ©Micronet S.A. 1999/2000
En resumen, que el ser humano se ha adaptado a diferentes ambientes y les ha acompañado a la invención de nuevas técnicas de supervivencia una serie de modificaciones morfológicas, que no son válidas para distinguir unas poblaciones humanas de otras desde un punto biológico. Además el color claro de la piel no es exclusiva de los europeos, ni los caucasoides son todos de piel blanca.
Lo de que el neandertal era de piel blanca y hasta rubio o pelirrojo, ya lo había oido hace tiempo, y me pareció curioso, pues tradicionalmente al neandertal siempre se le relacionó con un fenotipo moreno que recordaba al mediterraneo, considerado antiguamente más primitivo que el tipo nórdico.
El problema no es sólo o al menos no exclusivamente la existencia o no de determinados test realizados sobre el ADN, sea este mt o nuclear. Esto es una herramienta precisa...pero ¿podemos olvidar de golpe y porrazo las evidencias arqueológicas?.
Bien miradas, excepción hecha de la tipología del retoque y de algunas morfometrías, las industrias neanderthales y las sapiens son parejas. Es más, dentro de la concepción simbólica o conceptual del útil, existe la misma intencionalidad y la persecución de una finalidad y funcionalidad similares. ¡¡Hasta en las cadenas operativas se observan gestos técnicos similares!!. Pero podemos ir más lejos. Las evidencias de tratamiento simbólico-mágico dentro del concepto funerario permiten también ese acercamiento. Incluso la tipología de hábitats coincide en numerosas ocasiones...
Visto de forma tranquila, y obviando por un moento el ADN -al menos los estudios realizados hasta la fecha- ¿qué podemos concluir?. Aún más, ¿es, con el número de muestras tomadas, posible establecer una mínima fiabilidad estadística a los muestreos?. ¿es posible con los datos en la mano elAborar hipótesis sustentadas por varios cientos o miles de estadígrafos que avalen una u otra teoría?.
Respecto a lo de los dientes, y con todo el respeto para Bermúdez de Castro...¡¡¡por dios!!!, que él mismo codirige algún yacimiento en el que la datación se basa en dos míseras muelas. Es hora de que la arqueología toque ciencia y que hasta que no tengamos pruebas fehacientes y científicamente demuestrables y repetibles que de veras especifiquen algo, en vez de discutir, o vender humo, nos dediquemos a buscar datos, desenterrarlos con cautela y con los especialistas precisos, obetner muestreos amplios y buscar explicaciones globales que vayan más allá de la mera curiosidad de si un macho neanderthal y una hembra sapiens pudieron o no procrear...¡¡¡pero si hasta a algunos amigos mís -que son sapiens los dos, que yo sepa- les cuesta una babaridad tener un hijo....!!!!
A mi me parece aladelta que la cuestión de el cuestionamiento de la validez del término "raza" responde más a que la susodicha palabreja tiene peor prensa que el príncipe Carlos, y queda más guapo quitárselo de encima por sus connotaciones perversas, que desde luego no son cosa intrínseca del término sino que vienen del uso que se le ha dado en muchos episodios lúgubres de la historia humana.
¿Por qué está superado el término raza, porque la genética ha descubierto que somos mucho más cercanos genéticamente de lo que creíamos? ¿porque atendiendo a cierto marcador (cromosoma Y, ADNmt,...) un señor de una "etnia" puede estar más relacionado (en lo referente a ese marcador, claro) con los de otra "etnia" que con los de la suya?
¿cuál es exactamente la distancia genética requerida entre los individuos de dos poblaciones para poder empezar a hablar de razas? ¿hay algún acuerdo sobre eso?
Si en una pareja de bantúes el padre es del haplogrupo E (el mismo que comparten muchos europeos de aspecto totalmente caucasoide) y tienen un niño... ¿crees que hay alguna posibilidad de que el niño pase poe europeo caucasoide? ¿Cómo se puede negar pues la base genética de las diferencias entre poblaciones humanas que han mantenido entre sí un cierto aislamiento? Me l'expliquen.
Cambiar "raza" por "etnia" no afecta a más cuestiones que la puramente terminológica.
Afortunadamente, ni todos los antropólogos ni todos los genetistas se dejan llevar por la ola samaritana, corriendo el inevitable riesgo de ser clasificados como científicos "racistas".
En http://biblo.tech.nu/ hay unos cuantos artículos que Abordan el circo de la raza.
""" El titulo de este ensayo - sin las interrogantes - apareció en Internet de manera bastante frecuente a mediados del mes de Diciembre de 2002, como publicidad a un estudio genético realizado por varios investigadores brasileños (Parra et. al., "Color y ancestro genomico en los brasileños," PNAS 100: 177-182, 2003):
...
Los autores al poco tiempo hacen el siguiente comentario:
"Hay un gran acuerdo entre los antropólogos y los genetistas, que desde un punto de vista biológico, las razas humanas no existen."
¿De que manera este "gran acuerdo" coincide con los últimos estudios que muestran que las categorías raciales y étnicas son validas dado que los miembros de diferentes "grupos poblacionales" varían en la prevalencia de ciertas enfermedades, en la respuesta a los tratamientos médicos, y en la habilidad para aceptar trasplantes de medula de otros individuos pertenecientes a otros "grupos de poblaciones?""""
(Rienzi, M., "¿Los científicos comprueban que la raza no existe?")
La cuestión es que cuando los latinoamericanos describen a los españoles como "monos" o cuando los los ingleses piensan que todos somos morenos, falsean la realidad y generalizan (el todo por una parte), pues tales apreciaciones son eso, estereotipos sin el más mínimo raciocinio mediante. Pero si a tí te parece que es posible que haya un solo Aborigen australiano que pase por chino han, o viceversa, y que pensar lo contrario es cosa de cultura o de tópicos o de estar acostumbrado a tal o cual, en fin, yo qué quieres que te diga. Lo que yo creo es que un extraterrestre medianamente observador llegado ayer a la Tierra ya sabría hoy al mediodía clasificar sistemáticamente a un han como tal y a un australiano como tal (sin meterme en otras poblaciones del SE asiático o del Pacífico).
Evidentemente no todos los bantúes tienen la piel exactamente de la misma tonalidad, y evidentemente no todos los rasgos que diferencian a un grupo humano A de otro B, no comunicados, presentan la misma intensidad o el mismo desarrollo en todos los miembros del grupo en cuestión, pero llega un momento en la evolución por separado de ambos grupos en que la diferenciación es tal, que es imposible que el un miembro de A sea confundido con otro de B. Después, dependiendo de la extensión de los miembros de A, pueden aparecer subgrupos que desarrollen otros aspectos, subrazas, que si no se comunican entre sí, terminará ocurriendo lo que antes entre A y B.
Y es que las diferencias entre especies, razas, y en general grupos lo suficientemente definidos y diferenciados para clasificarlos como distintos, es siempre CUALITATIVA, y no CUANTITATIVA. Pero claro, la cuestión es que, cuantitativamente, llega un momento en que la diferencia es tal que con ver a un individuo sabrás sin género de dudas a qué grupo pertenece, pues el rasgo (o rasgos) que lo carazterizan como perteneciente a ese grupo, se inscribe en un arco de posibilidades del cual están excluidos todos los individuos del otro grupo. Seguramente no todos los neanderthales eran de la misma estatura, ni todos tenían exactamente la misma capacidad craneal, ni el arco supracilial desarrollado en la misma forma, ni la misma complexión, seguramente había variedad de rasgos en lo que llamamos homo neanderthalensis. Lo mismo para nuestra especie. Y lo más importante: los sapiens y los neanderthales no surgieron del suelo como setas, fueron evolucionando lentamente a partir de un tronco común, lo que quiere decir que por fuerza hubo poblaciones con rasgos intermedios, en toda la escala gradual, entre sapiens y neandertales, y quien sabe si contemporáneas a los propios sapiens y neanderthales.
¿Donde empieza una especie y donde acaba la otra? Qué, ¿tiramos el concepto de especie a la basura? Como verás, la diferencia entre especies también es cualitativa. ¿Donde acaba la raza y donde empieza la especie? Interesantes cuestiones, no? El problema, y lo que tenemos que tener presente, es que raza y especie son constructos humanos, que tienen su realidad en nuestra mente y que son una valiosa herramienta para procesar la información. Que no coincida exactamente con la realidad material,... ¿quiere decir que hemos de desecharlos?
La dferencia entre frío y caliente es gradual. ¿No son válidos pues los conceptos "frío" y "caliente"? También es gradual la diferencia entre rojo y naranja, entre un genio y un tonto, entre sofá y silla (porque siempre habrá aparatos intermedios), entre homínido y no homínido (donde empiezan los homínidos?)... continuamente, en la vida cotidiana como en el campo científico, realizamos clasificaciones de este tipo.
¿Por qué vale "especie" y no "raza"? Yo te lo diré:
a) por la mala prensa de "raza" y la bondad samaritana de nuestros bienpensantes
b) porque los neanderthales, extintos ya los pobres, no votan y no se quejan. Si siguieran conviviendo con nosotros, me imagino que ya habrían surgido voces cuestionando la conveniencia de considerarlos otra "especie"... jojojo, me troncho (fíjate, a lo mejor hasta resurgía el interés por el concepto de "raza")
Debería de leer los mensajes que escribo antes de enviarlos. He decidido repetirlo aquí entero, corregido, para que su lectura no resulte un lío:
La cuestión es que cuando los latinoamericanos describen a los españoles como "monos" o cuando los los ingleses piensan que todos somos morenos, falsean la realidad y generalizan (el todo por una parte), pues tales apreciaciones son eso, estereotipos sin el más mínimo raciocinio mediante. Pero si a tí te parece que es posible que haya un solo Aborigen australiano que pase por chino han, o viceversa, y que pensar lo contrario es cosa de cultura o de tópicos o de estar acostumbrado a tal o cual, en fin, yo qué quieres que te diga. Lo que yo creo es que un extraterrestre medianamente observador llegado ayer a la Tierra ya sabría hoy al mediodía clasificar sistemáticamente a un han como tal y a un australiano como tal (sin meterme en otras poblaciones del SE asiático o del Pacífico).
Evidentemente no todos los bantúes tienen la piel exactamente de la misma tonalidad, y evidentemente no todos los rasgos que diferencian a un grupo humano A de otro B, no comunicados, presentan la misma intensidad o el mismo desarrollo en todos los miembros del grupo en cuestión, pero llega un momento en la evolución por separado de ambos grupos en que la diferenciación es tal, que es imposible que el un miembro de A sea confundido con otro de B. Después, dependiendo de la extensión de los miembros de A, pueden aparecer subgrupos que desarrollen otros aspectos, subrazas, que si no se comunican entre sí, terminará ocurriendo lo que antes entre A y B.
Y es que las diferencias entre especies, razas, y en general grupos lo suficientemente definidos y diferenciados para clasificarlos como distintos, es siempre CUANTITATIVA, y no CUALITATIVA. Pero claro, la cuestión es que, cuantitativamente, llega un momento en que la diferencia es tal que con ver a un individuo sabrás sin género de dudas a qué grupo pertenece, pues el rasgo (o rasgos) que lo carazterizan como perteneciente a ese grupo, se inscribe en un arco de posibilidades del cual están excluidos todos los individuos del otro grupo. Seguramente no todos los neanderthales eran de la misma estatura, ni todos tenían exactamente la misma capacidad craneal, ni el arco supraorbital desarrollado en la misma forma, ni la misma complexión, seguramente había variedad de rasgos en lo que llamamos homo neanderthalensis. Lo mismo para nuestra especie. Y lo más importante: los sapiens y los neanderthales no surgieron del suelo como setas, fueron evolucionando lentamente a partir de un tronco común, lo que quiere decir que por fuerza hubo poblaciones con rasgos intermedios, en toda la escala gradual, entre sapiens y neandertales, y quien sabe si contemporáneas a los propios sapiens y neanderthales.
¿Donde empieza una especie y donde acaba la otra? Qué, ¿tiramos el concepto de especie a la basura? Como verás, la diferencia entre especies también es cuantitativa. ¿Donde acaba la raza y donde empieza la especie? Interesantes cuestiones, no? El problema, y lo que tenemos que tener presente, es que raza y especie son constructos humanos, que tienen su realidad en nuestra mente y que son una valiosa herramienta para procesar la información. Que no coincida exactamente con la realidad material,... ¿quiere decir que hemos de desecharlos?
La dferencia entre frío y caliente es gradual. ¿No son válidos pues los conceptos "frío" y "caliente"? También es gradual la diferencia entre rojo y naranja, entre un genio y un tonto, entre sofá y silla (porque siempre habrá aparatos intermedios), entre homínido y no homínido (donde empiezan los homínidos?)... continuamente, en la vida cotidiana como en el campo científico, realizamos clasificaciones de este tipo.
¿Por qué vale "especie" y no "raza"? Yo te lo diré:
a) por la mala prensa de "raza" y la bondad samaritana de nuestros bienpensantes
b) porque los neanderthales, extintos ya los pobres, no votan y no se quejan. Si siguieran conviviendo con nosotros, me imagino que ya habrían surgido voces cuestionando la conveniencia de considerarlos otra "especie"... jojojo, me troncho (fíjate, a lo mejor hasta resurgía el interés por el concepto de "raza")
PROYECTO GENOGRAPHIC
National Geographic e IBM inician un estudio genético mundial para rastrear
Nacional Geographic Society e IBM han anunciado hoy un proyecto antropológico a escala mundial para recopilar una de las bases de datos de genética más grandes del planeta. El objetivo es rastrear el proceso de población de la Tierra a lo largo de la Historia. El proyecto pretende revelar detalles sobre la historia migratoria de la humanidad y contribuir al conocimiento sobre las similitudes y diferencias que marcan la especie humana.
12:09 - 14/04/2005 | Fuente: REDACCIÓN, LAFLECHA
El proyecto, denominado Genographic y cuya duración está prevista para cinco años, consiste en el análisis en lAboratorio y mediante el uso de la informática de ADN obtenido de pueblos indígenas y donado por cientos de miles de personas. Un equipo de científicos de National Geographic y de IBM, dirigidos por el investigador Spencer Wells, recopilará las muestras de ADN alrededor del mundo y analizará los resultados para establecer las raíces genéticas de los humanos modernos. Con financiación de la ?Waitt Family Foundation?, los científicos establecerán 10 centros en todo el mundo para estudiar más de 100.000 muestras de ADN.
?Vemos este proyecto como un paso de gigante en antropología, al permitirnos utilizar la genética para rellenar los huecos en nuestro conocimiento de la historia humana?, explica Spencer Wells, director del proyecto. ?Nuestro ADN contiene una historia que compartimos todos. Durante los próximos cinco años estaremos descifrando esa historia, que ahora corremos el riesgo de perder porque la gente emigra y se mezcla a un ritmo mucho más rápido que en el pasado?.
Los ciudadanos de todo el mundo pueden participar en el proyecto Genograhic permitiendo que sus datos de ADN se incluyan en la base de datos. Los individuos podrán seguir el progreso de su propia historia migratoria y de la investigación global conectándose a la pagina web de National Geographic (www.nationalgeographic.com). La base de datos resultante será una de las mayores recopilaciones de datos genéticos del mundo y servirá como recurso para genetistas, historiadores y antropólogos.
Científicos del Centro de Biología Computacional de IBM, una de las instalaciones punteras en investigación biocientífica, utilizarán tecnologías de análisis avanzado y de clasificación de datos para interpretar las muestras y descubrir nuevos patrones y conexiones en los datos que contienen. IBM también proporcionará el conocimiento informático y la infraestructura para la gestión de los cientos de miles de códigos genéticos que serán analizados por el proyecto Genographic.
?National Geographic ha explorado y cartografiado el mundo desde hace 117 años?, comenta John Fahey, presidente de National Geographic Society. "Ahora, como resultado de esta asociación con IBM, estamos desplegando la ciencia y tecnología más modernas para establecer nuestro viaje por todo el planeta. Esperamos que este ambicioso proyecto incremente nuestro
entendimiento y conocimiento de la Historia. El trabajo de campo de esta investigación formará parte de un museo virtual de la Historia Humana?.
El proyecto Genographic tiene tres componentes clave:
Trabajo de campo: la recolección de muestras de ADN de pueblos indígenas y el trabajo de investigación de campo con este tipo de poblaciones son una de las claves del proyecto. Las muestras de sangre de poblaciones indígenas, cuyo ADN contiene datos que han permanecido relativamente inalterados durante cientos de generaciones, son indicadores fiables de patrones migratorios antiguos. Spencer Wells y un consorcio de científicos de instituciones internacionales se encargarán de la recolección de datos y el trabajo de lAboratorio. Un consejo asesor internacional supervisará la selección de poblaciones indígenas y el seguimiento estricto de protocolos de investigación determinados.
Participación pública y de campaña de concienciación: los ciudadanos de todo el mundo pueden participar comprando un ?kit de participación? por 99,95 dólares y enviando sus propias muestras de células (del interior de la boca). Ello les permite hacer un seguimiento de todo el proyecto y de su propia historia migratoria. Estos resultados personales se almacenarán de forma segura y anónima para asegurar la privacidad de los participantes. National Geographic e IBM informarán periódicamente a la comunidad científica sobre los progresos de la investigación a través de un sitio en Internet y otros medios de los que dispone National Geographic en todo el mundo. Un programa de televisión, ?En Busca de Adán?, será emitido en Estados Unidos y por todo el mundo a través de National Geographic Channel?.
El legado del proyecto Genographic: los recursos obtenidos por la venta de los kits de participación servirán para financiar el futuro trabajo de campo y para apoyar proyectos de preservación cultural y soporte a la educación entre las poblaciones indígenas que hayan participado.
http://www.laflecha.net/canales/ciencia/noticias/200504132/
Por qué sesgada por que las mouestras las recojan sólo personas con estudios superiores? Y en cuanto a que se reduzca al hemisferio norte, hombre, los de NG tienen colAboradores por todo el planeta, y es lógico pensar que si quieren hacer un buen trabajo, algo interesante, habrán de incidir especialmenet en minorías y grupos apartados.
Mira, en la página de NG dicen:
"The Legacy Project will provide indigenous groups participating in the Genographic Project with direct help through development projects, education, and public-awareness campaigns aimed at preserving traditional cultures."
Más noticias sobre el homo floresiensis:
La paleoneuróloga Dean Falk, la máxima responsable del departamento de antropología de la Universidad del Estado de Florida, llega a la conclusión de que los restos del “hobbit” encontrado en la cueva Liang Bua son de un homínido diferenciado: el homo floresiensis.
Empezó examinando el cráneo por si fuera cierto que pertenecía a un individuo con microcefalia, según cree el arqueólogo Teuku Jacob; ella encontró grandes diferencias entre un cráneo de un individuo que sufría microcefalia y que comparó con el del famoso hobbit; concluyó que la forma del cráneo de un microcefálico difiere bastante del cráneo del hobbit, y que por tanto el del “hobbit” no era un cráneo microcefálico. Luego comparó el cerebro de un pigmeo con el reconstruido a partir del cráneo del “hobbit”, y vio que el del pigmeo era un cráneo enorme al lado del “hobbit” siendo tres veces más grande. Con lo que llegó a la conclusión de que el “hobbit” no era un pigmeo. Se empezaron, pues, a compararlo con cerebros de chimpancés y otros primates, y, por supuesto, con otros homínidos del registro fósil, y se dieron cuenta que el candidato al que más se parecía era el del el cerebro de un homo erectus. Mirado por cada ángulo, el cerebro reconstruido del “hobbit” es muy parecido en forma (que no en tamaño) al del homo erectus.
Pero la siguiente pregunta que nos hacemos todos es ¿cómo pudo retener la inteligencia de su antepasado erectus (hacer fuego, cazar, e incluso, como dicen las leyendas de los aldeanos locales, tener un tipo de lenguaje con una capacidad craneana de apenas 300 cc?
La respuesta parece estar en la parte frontal del cerebro. La llamada Área 10. Todos los simios grandes y seres humanos tienen éste área 10, pero a diferencia de nosotros el “hobbit” la tiene excepcionalmente grande. El Area 10 es la responsable de muchos procesos de los cuales nos caracterizamos los humanos, como planear algo para el presente o futuro, tomar la iniciativa, llevar a cAbo cualquier intención. Pues bien, el área 10 del cerebro del “hobbit” es mayor que la del homo erectus; es posible según los científicos que mayor desarrollo del área 10 en el cerebro del homo floresiensis haya compensado el encogimiento del cerebro del antepasado del “hobbit”, reteniendo así la inteligencia del erectus en un cerebro más de dos veces pequeño.
Cuando los lugareños describen a los míticos Ebu Gogo, ésta descripción coincide con la de la reconstrucción efectuada por un lAboratorio del homo floresiensis.
Sacado del documental: Riddle of the Human Hobbits.
Sitios en internet:
http://news.telegraph.co.uk/news/main.jhtml?xml=/news/2005/03/04/whobb04.xml
http://www.stevequayle.com/News.alert/04_Unexplained/041028.hobbits.html
Un sitio en castellano:
http://usuarios.lycos.es/criptozoo/Hombrecillos/Hombrecillos.html
El homo erectus: ¿un antepasado MUY cercano?:
El erectus fue el primer hombre que salió de África para poblar el sur de Asia y presumiblemente Europa. Usaba piedras, preferiblemente cantos rodados de tamaño suficiente como para mantener a raya sus depredadores. El erectus se caracteriza por ser el primer hombre que intenta dominar el medio en el que vive.
Un fósil encontrado demuestra que el erectus cuidaba de los suyos, como lo haría un hombre moderno. Parece que pertenecía a un erectus que sufría de hinchazón ósea provocada probablemente por un exceso de vitamina A seguramente por excesivo consumo de hígado; de alguna manera sobrevivió largo tiempo con esta condición, y todo apunta a que otro miembro de su grupo habría cuidado del compañero enfermo.
El erectus fue también el primero en saber hacer fuego. Antes del fuego sólo se podía huir de los depredadores subido a un árbol, o manteniéndose lo más lejos posible de ellos; después de conocer el fuego, pudo permanecer en el suelo, ya que el fuego mantiene alejados a los animales.
Científicos que han observado a los chimpancés han llegado a la conclusión de que a éstos les encanta la carne y aprovechan cualquier oportunidad para matar monos más pequeños para devorarlos. Sin embargo la carne cruda es difícil de mascar por lo que se ha calculado que un chimpancé consigue las mismas calorías por hora comiendo carne que comiendo fruta. Por o tanto la carne no fue un alimento importante para los primeros humanos hasta que descubrieron cómo hacerla más tierna y fácil de mascar. El fuego ahorra unas 5 horas de mascado al día, reduciéndola a sólo 1 hora. Las calorías rápidas de comida cocinada provocó que el erectus fuera un 50% más alto que sus antecesores, alcanzando incluso nuestra propia altura; además la comida cocinada requiere menos digestión, por lo que sus intestinos y la caja torácica se hicieron más pequeños; gracias a una comida más tierna, los músculos de la mandíbula del erectus encoge, liberando el cráneo y el cerebro para ser más grande. Esto hizo al erectus/ergaster nuestro ancestro directo. De echo podría haberse alimentado de otro humano contemporáneo suyo, el Paranthropus Boisei, que desapareció sin tener descendencia hace 1’2 millones de años.
El heidelbergensis es descendiente del erectus/ergaster. Su cuerpo consumía el doble de energía que los nuestros. Su enorme tamaño indica que vivió en lugares donde había abundante caza de animales grandes. Su principal problema: mantenerse refrigerado; su corpulencia y metAbolismo rápido le hacía estar limitado a ambientes secos cercanos a ríos o fuentes de agua pues requeriría mucho consumo de ésta. Se cree que 500.000 años atrás éste fue el segundo humano que salió de Africa, después del erectus, y desapareció hace unos 240.000 años dando lugar a un descendiente más delgado y grácil (nosotros) y a otro igual de robusto pero más rechoncho (el neandertal).
Aunque aparentemente el trabajo-recopilación sobre genética fue realizado en 2005, no fue hasta hace pocos días, cuando me encontré con este artículo de forma casual, buscando algo de información sobre genética, mas que nada por distraerme. La sorpresa fue que mientras leía la interesante recopilación, realizada por “Aladelta” en un resumen sobre un trabajo sobre el cromosoma Y realizado por la universidad de Stanford con varios colAboradores de diferentes países, aparecía un dato que me parece erróneo. En la recopilación ó resumen, no en el estudio, ya que lo he descargado de la página original, decía que en Galicia, el 46% de la población tenía linaje africano. Creo que se ha equivocado al sumar, ya que sumando los haplogrupos de origen africano, que según lo que he podido entender son los E3a, E3b, etc (5,3+5,3+10,5+10,5=31,6% y no 46%) De hecho si el haplogrupo R1 es 63,1% (uno de los más elevados) el restante nunca llegaría ni siquiera al 40%.
El caso de Málaga que cita, si que parece correcto, aunque sólo sea matemáticamente, ya que 3,8+7,7+11,5+3,8+3,8=30,6% .
Bueno, hasta aquí todo casi normal, ya que un error lo puede tener cualquiera, pero vamos a centrarnos en el estudio en sí.
Me he fijado en el caso de Galicia y Málaga porque son los que tienen los índices más altos de linaje teóricamente africano, ¡vaya, que casualidad! Galicia más población con “sangre” africana que una provincia de Andalucía, vamos, de risa, resulta que 800 años de asentamiento de la cultura árabe, y tiene menos linaje africano que Galicia, que apenas sufrió incursiones militares ó razzias esporádicas.(Por cierto, ya quisiera Galicia haber tenido asentada en su territorio la espléndida civilización musulmana). Pero vamos con el método; según creo entender se han sometido a pruebas, por ejemplo para Cataluña 16 personas, Galicia 19 personas, cantidades totalmente ridículas, sin embargo en Sevilla 155. ¿Por qué estas diferencias?¿Qué eran, voluntarios, un grupo de amigos, ó los escogieron por la calle según su aspecto? ó hay otras motivaciones, incluso políticas, ya que estos estudios si no se dan en poblaciones al azar están totalmente invalidados. También es de extrañar la ausencia total de población con linaje africano en Cataluña. Cero. Vamos, hombre, esto no se lo cree nadie, cualquiera que haya visitado Cataluña, región mediterránea hasta la médula, (y con orgullo) puede comprobar que hay mucha variedad de tipos humanos, en fin, que no es la islas feroe,(por fortuna), que todos son rubios y comen bacalao.
No me extraña que luego escriban artículos en esta web, gente como Arkaitz2, que dijo literalmente:9-12-2003 “La verdad es que yo, quizá debido a mi ignorancia, no veo actualmente ningún sustrato celta en los gallegos: hablan una lengua romance, su tez es tan oscura como la de un habitante de la cuenca mediterránea, sus hábitos son básicamente peninsulares. ¿Qué queda de celta? ¿No hay más sustrato árabe en ellos que celta? ¿Por qué toda la gente en esta página se empeña en ver celtas puros por todas partes? Para bien o para mal, lo celta no fue la esencia de ningún pueblo peninsular y con el tiempo se ha convertido en pura anecdota.”
Vamos, que en el país vasco no hay morenos, según él, y también cree que la piel del resto de pueblos ibéricos es “oscura”, bueno, es tan oscura como la de los vascos, se nota que no ha viajado el pobre, ni dentro del país vasco siquiera. Yo conozco a vascos de más de 8 apellidos vascos y tienen la piel tan morena como un habitante de la cuenca mediterránea de los más oscuritos. Peor que un centralista radical antinacionalista, es un nacionalista que quiere ser de primera y niega y desprecia a los pretendidamente de segunda, poniéndose la venda en los ojos y negando obviedades como la inmensa toponimia celta en Galicia y Asturias, sin olvidar a los que ven celtas y celtismo por todas partes, y tampoco es eso,
También me he fijado que colAboran científicos españoles, vamos que cada uno hace un estudio sobre su país, o incluso sobre su región, con la falta de “asepsia “ que esto conlleva y se lo mandan a Stanford, y allí ellos, con sus batitas blancas, se ríen de nosotros, y se dicen entre sí, pero que gilipollas son estos españoles.
Como conclusión, creo que si se hacen estudios genéticos, se deberían hacer con más seriedad y aplicando el método científico, sino crean confusión, desconfianza, y siempre hay interesados y tergiversadores, cuando no partidarios de la eugenesia.
dasagis06 de abr. 2008Información eliminada por el Administrador Silberius
dasagis11 de abr. 2008Información eliminada por el Administrador Silberius
dasagis11 de abr. 2008Información eliminada por el Administrador Silberius
Hay 21 comentarios.
1